تکثیر سلولهای بنیادی خونساز CD34+ بند ناف در همکشتی با سلولهای بنیادی مزانشیمی مغز استخوان در حضور TEPA
ناصر امیریزاده1، فرهاد ذاکر2، ناهید نصیری3
چکیده سابقه و هدف استفاده از سلولهای بنیادی خونساز(HSCs)، روش استانداردی جهت درمان بسیاری از بدخیمیهای خونی و اختلالات غیرخونی محسوب میشود. خون بند ناف، دارای مزایای فراوانی نسبت به سایر منابع سلولهای بنیادی است و یکی از محدودیتهای مهم این منبع جهت پیوند, تعداد محدود سلولهای بنیادی آن میباشد. از راهکارهای پیشنهادی برای غلبه بر این مشکل، تکثیر بدون تمایز این سلولها در محیطهای کشت است. در این مطالعه از TEPA به عنوان شلاتور مس و سلولهای بنیادی مزانشیمی مغز استخوان جهت تکثیر UCB-HSCs استفاده شد. مواد و روشها مطالعه انجام شده از نوع کاربردی بود. سلولهای CD34+ خون بند ناف با استفاده از آنتیبادیهای نشاندار با Microbead و ستون MACS جدا شدند. سپس این سلولها در چهار شرایط کشت مختلف شامل: 1) محیط حاوی سایتوکاین 2) محیط دارای فیدر و سایتوکاین 3) محیط دارای TEPA و سایتوکاین و 4) محیط دارای فیدر, سایتوکاین و TEPA کشت داده شدند. به منظور ارزیابی تکثیر HSCs، از شمارش سلولی, تعیین درصد سلولهای CD34+ به روش فلوسایتومتری و روش سنجش کلونی در روز 10 تکثیر استفاده شد. یافتهها بیشترین میزان چند برابر شدن تعداد سلولهای CD34+, TNCها و CFU-C در شرایط کشت چهارم(به ترتیب 3/15 ± 11/110, 78/21 ± 5/118 و 7/44 ±9/172 برابر) در مقایسه با سایر شرایط مشاهده شد. نتیجه گیری نتایج بررسی نشان داد که همکشتی HSCs با سلولهای بنیادی مزانشیمی در حضور عامل شلاتهکننده مس ((TEPA، قادر است میزان تکثیر UCB-HSCs را به طور قابل توجهی افزایش دهد. بنابراین، این استراتژی میتواند برای تکثیر HSCs مفید واقع شود. کلمات کلیدی: سلولهای بنیادی خونساز, سلولهای بنیادی مزانشیمی, TEPA
تاریخ دریافت : 4/11/91 تاریخ پذیرش : 21/1/92
1- PhD هماتولوژی و بانک خون ـ استادیار مرکز تحقیقات انتقال خون ـ مؤسسه عالی آموزشی و پژوهشی طب انتقال خون ـ تهران ـ ایران 2- PhD هماتولوژی ـ استاد دانشکده پیراپزشکی ـ مرکز تحقیقات سلولی و مولکولی دانشگاه علوم پزشکی ایران ـ تهران ـ ایران 3- مؤلف مسؤول: کارشناس ارشد هماتولوژی ـ دانشکده پیراپزشکی دانشگاه علوم پزشکی تهران ـ تهران ـ ایران ـ صندوق پستی: 6183-14155
مقدمه پیوند سلولهای بنیادی طی دهههای اخیر به موفقیتهای درمانی بالایی رسیده است و یک راهکار درمانی مناسب و عملی برای بسیاری از بیماریهایی که در گذشته غیر قابل درمان بودند را به وجود آورده است(3-1). ویژگیهای اصلی سلولهای بنیادی خونساز(HSCs) ، پتانسیل خود نوسازی(selfrenewality) و تمایز به انواع مختلف سلولهای خونی میباشد(5، 4). خون بند ناف(UCB) به دلیل بالا بودن تعداد اهداکنندگان، دسترسی راحت و شیوع کمتر GVHD (بیماری پیوند علیه میزبان) علیرغم عدم سازگاری کامل HLA، جایگزین مناسبی برای مغز استخوان جهت پیوند سلولهای بنیادی خونساز میباشد(6). صرف نظر از این مزایا، محدودیت عمده خون بند ناف تعداد کم HSCs موجود در آن جهت پیوند به بیماران بزرگسال است. سرعت پیوند به تعداد سلولهای بنیادی CD34+ خون بند ناف بستگی دارد. دوز مطلوب جهت انجام پیوند در بزرگسالان بیشتر یا مساوی CD34+ cell/kg 106 × 5/2 میباشد. اگر نمونه خون بند ناف به صورت مناسب جمعآوری شود، دارای 107 سلول CD34+ خواهد بود که این تعداد تنها جهت استفاده در یک نوزاد کافی میباشد(12-7). راهکار پیشنهادی که امروزه بسیار مورد توجه قرار گرفته است، تکثیر HSCs در محیطهای ex vivo میباشد. میتوان این سلولها را در حضور سایتوکاینها در محیط کشت تکثیر نمود اما متاسفانه تکثیر HSCs در آزمایشگاه با مشکلاتی مواجه است زیرا HSCs در in vitro به تدریج توانایی خودسازی و چند قوهای بودن خود را از دست داده و به ردههای خاصی تمایز مییابند(14، 13). احتمالاً تمایز سلولها در شرایط in vitro ناشی از عدم وجود تنظیماتی است که در in vivo توسط ریز محیط اعمال میشود. در مغز استخوان HSCs عمدتاً در تعامل با ریز محیط ویژهای به نام نیچ قرار دارند که سرنوشت سلولها را از نظر خاموشی(quiescence), خودنوسازی و تمایز تنظیم میکند. مطالعههای اخیر نشان دادهاند که تماس مستقیم بین سلولهای استرومال و HSCs ، برای بقا، تنظیم سیکل سلولی، مهاجرت و پیوند مهم میباشد. در بسیاری از مطالعهها، اثرات سینرژیستیکی با استفاده از سایتوکاینهای نوترکیب و لایه فیدر مشتق از سلولهای بنیادی مزانشیمی (MSCs)در تکثیر و تمایز HSCs گزارش شده است(21-15). راهکار دیگری که اخیراً جهت تکثیر بدون تمایز سلولهای بنیادی خونساز در شرایط in vitro به کار گرفته شده است، استفاده از شلاتور مس(TEPA) همراه با سایتوکاینهای Early acting از جمله SCF ، TPO و FL جهت ازدیاد تکثیر سلولهای پیشساز خونساز(HPCs) و سلولهای بنیادی خونساز(HSCs) بند ناف میباشد. از آن جایی که میزان بالای مس درون سلولی به دلیل نقش آن در تولید رادیکالهای آزاد اکسیژن توکسیک است، کاهش میزان این رادیکالها جهت حفظ HSCs در مغز استخوان و اریتروپوئز طبیعی ضروری است. عمده مس سلولی به اجزای سلولی از جمله سرولوپلاسمین و آنزیمهای مختلف متصل است و تنها مقدار کمی از آن به فرم یونیزه و آزاد وجود دارد. فرم یونیزه مس قابلیت اتصال به شلاتورها را دارد که به آن Chelatable CU میگویند(27-22). بدین منظور در این مطالعه تاثیر شلاتور مس بر میزان تکثیر سلولهای CD34+ بند ناف در همکشتی با سلولهای بنیادی مزانشیمی مغز استخوان, به عنوان لایه فیدر, در شرایط مختلف کشت بررسی و مقایسه شد.
مواد و روشها جداسازی سلولهای CD34+ : در این مطالعه کاربردی، سلولها از خون بند ناف پس از دریافت فرم رضایتنامه جدا شدند. ابتدا سلولهای تک هستهای(MNCs) مربوط به 5 نمونه خون بند ناف به کمک فایکول(سیگما، kg/L 001/0 ± 077/1) جدا شد. سپس این سلولها با آنتی CD34 کونژوگه شده با ذرات میکروبید(آمریکا، میلتنی بیوتک) انکوبه گردیده و در نهایت با استفاده از روش MACS (آمریکا، میلتنی بیوتک)، سلولهای CD34+ موجود در بین MNCها با دو بار عبور از ستون LS از سایر سلولها جدا شدند .در نهایت جهت ارزیابی درجه خلوص سلولهای CD34+ جدا شده، آنالیز فلوسایتومتری با استفاده از آنتیبادیهای آنتی CD34 (آمریکا، بیوساینس) و آنتی CD38 انجام شد. میانگین متوسط سلولهای CD34+ حدود 62% بود(28).
جداسازی و کشت سلولهای بنیادی مزانشیمی: MSCs از نمونه آسپیره مغز استخوان اهداکنندگان سالم در ضد انعقاد هپارین جداسازی شد(رضایتنامه از آنها گرفته شد). سلولهای تک هستهای مغز استخوان به وسیله گرادیان شیب غلظت فایکول(سیگما، kg/L 001/0 ± 077/1) جدا شده و پس از شستشو در محیط DMEM (آمریکا، سیگما) Low glucose ، حاوی 10% FBS کشت داده شدند. پس از 48 ساعت انکوباسیون، سلولهای غیر چسبنده با تعویض محیط خارج گردیدند. پس از 3 پاساژ، سلولهای چسبنده مزانشیمی از محیط قابل برداشت بودند. فنوتیپ MSCs با استفاده از آنتیبادیهای (Copenhagen ، دانمارک، داکو) آنتی CD45 ، CD37 ، CD166 ، CD105 ، CD90 و CD34 توسط فلوسایتومتری مورد ارزیابی قرار گرفت. سپس 104 * 1 سلول مزانشیمی در پلیتهای 6 خانه(Roskilde ، دانمارک و Nunc) کشت داده شدند تا به 80% از پوشیدگی سطح رسیدند. به منظور جلوگیری از تکثیر بیشتر، سلولها با میتومایسین C تیمار شدند(19).
بررسی پتانسیل تمایزی سلولهای بنیادی مزانشیمی: هم چنین به منظور تایید این سلولها، تمایز آنها به سمت رده استئوبلاستی با استفاده از محیط تمایزی حاوی mM 10 دگزامتازون(آمریکا، کمیکون) ، M 1/0 آسکوربیک اسید ، DMEM-Low glucose (آمریکا، سیگما) و mol 1 گلیسرول دو فسفات(آمریکا، کمیکون) انجام شد. در نهایت سلولهای استئوبلاست با دو رنگآمیزی اختصاصی آلیزارین رد(آمریکا، سیگما) و آلکالن فسفاتاز(آمریکا، سیگما) مـورد تاییـد قرار گرفتند(29).
بررسی MTT: MTT (3-[4,5-dimethylthiazol-2-yl]-2, 5-diphenyl tetrazolium bromide) روشی است که فعالیت سلولهای زنده را از طریق دهیدروژنازهای میتوکندریایی مورد ارزیابی قرار میدهد. با استفاده از روش MTT ، اثرات سایتوتوکسیک احتمالی TEPA (آمریکا، سیگما) بر روی MSCs در محیط کشت، پس از 3 روز انکوباسیون تعیین شد. در هر یک از چاهکهای پلیت 96 خانهای بر روی MSCs ، رقتهای مختلف TEPA اضافه شد(هر رقت به صورت 3 تایی تکرار شد). سپس پلیتها در دمای 37 درجه سانتیگراد و فشار 5% CO2 انکوبه شدند. پس از اتمام زمان انکوباسیون، محیط کشتها کاملاً خالی گردیده و به هر چاهک محیط کشت جدید و محلول MTT اضافه شد. پس از اتمام زمان انکوباسیون، محیط کشت چاهکها تخلیه شده و با حل کردن بلورهای فورمازان در DMSO (دی متیل سولفوکساید)، جذب نوری نمونهها در طول موج 570 نانومتر با الایزا ریدر قرائت شد(30).
اندازهگیری یون مس: میزان مس سلولی طبق مطالعههای گذشته اندازهگیری شد. سلولها جمعآوری گردیده و با PBS شستشو داده شدند. 106 سلول در لوله اپندورف متال فری سانتریفیوژ گردید. پلیت سلولی با اسید نیتریک اولترا پیور (Netherlands ، Deventer ، Mallinckrodt Baker B.V) M 03/0 به غلظتmL / cell1010 * 1 رسانده شد. نمونهها سونیکیت شده و سپس با اتمیک ابزربشن کوره(ژاپن، شیمادزو، G 670 model) با طول موج 7/324 نانومتر اندازهگیری شدند(23).
کشت سلولهای CD34+ در شرایط مختلف: سلولهای CD34+ جدا شده در شرایط کشت مختلفی با غلظت cell/mL 104 * 1 به مدت 10 روز کشت داده شدند. این شرایط کشت عبارت بودند از: 1) شرایط کشت اول که سلولهای CD34+ جدا شده تنها در حضور سایتوکاینهای نوترکیب شامل؛ Stem Cell Factor (SCF) ، FLt3-Ligand (FL) وThrombopoietin (TPO) با غلظت ng/mL100 کشت داده شدند. جهت هر 4 شرایط کشت از همین سایتوکاینها با مشخصات ذکر شده و محیط کشت فاقد سرم(Stemspan) استفاده شد. 2) شرایـط کشـت دوم که سلولهای CD34+ جدا شده در همکشتی با MSCs به عنوان لایه فیدر و در حضور سایتوکاینها کشت داده شدند. پس از رسیدن MSCs به 80% از پوشیدگی سطح، محیط Stemspan حاوی ترکیب سایتوکاینی فوق به همراه 104 سلول CD34+ جدا شده، بر روی لایه فیدر سلولهای مزانشیمی کشت داده شدند. 3) شرایط کشت سوم که سلولهای CD34+ جدا شده در حضور سایتوکاینها و µM10TEPA کشت داده شدند. 4) کشت سلولها در محیط چهارم که سلولهای CD34+ جدا شده در همکشتی با MSCs به عنوان لایه فیدر و در حضور سایتوکاینها و µM10TEPA کشت داده شدند. تمامی محیط کشتها در انکوباتور 37 درجه سانتیگراد و 5% CO2 انکوبه شدند. تمامی کشتها به صورت سه تایی انجام شده و تعویض محیط در روزهای 3 , 7 و 10 صورت پذیرفت. قبل از آغاز کشت و پس از تکثیر، تعداد کل سلولهای هستهدار شمارش گردیده و سلولها با فلوسایتومتری جهت مارکر CD34 مورد ارزیابی قرار گرفتند.
سنجش کلونی(CFU assay) : 103 × 1 سلول از سلولهای CD34+ جدا شده از بند ناف و همچنین سلولهای پس از تکثیر در شرایط کشت مختلف در روز 10، با محیط methocult H4435 (استم سل تکنولوژی) مخلوط شده و درون پلیتهای 2mm 35 (دانمارک، Nunc ) ریخته شدند. این مرحله به صورت دوتایی انجام شد و پلیتها به مدت 14 روز در دمای 37 درجه سانتیگراد و 5% CO2 انکوبه شدند. پس از این مدت کلونیها به وسیله میکروسکوپ اینورت(ژاپن، نیکون) شمارش و بررسی شدند(32، 31).
آنالیز فلوسایتومتری: سلولها قبل(روز صفر) و بعد از تکثیر در شرایط کشت مختلف در روز 10 برداشته شده و با آنتیبادیهای مونوکلونال(کوپنهاگ،داکو، دانمارک) آنتی FITC-CD34 و PE-CD38 رنگآمیزی شدند. از آنتیبادی IgG موشی (کوپنهـاگ، دانمارک، داکو) به عنوان ایزوتایپ کنترل استفاده شد. در نهایت به وسیله دستگاه فلوسایتومتری(آلمان، پارتک) درصد سلولهای CD38-CD34+ ارزیابی شد(33).
آنالیز آماری: تجزیه و تحلیل آماری با استفاده از نرمافزار آماری 16SPSS انجام شد. جهت مقایسه گروههای مختلف سلولی و آنالیز نتایج از ANOVA استفاده گردید. 05/0 p< به عنوان معنادار در نظر گرفته شد.
یافتهها تایید سلولهای بنیادی مزانشیمی: ایمونوفنوتایپ این سلولها با استفاده از آنالیز فلوسایتومتری ارزیابی شد. بیان CD90 ، CD105 ، CD166 و CD37 به ترتیب 8/96% ، 93% ، 8/98% و 94% بود، در حالی که بیان CD34 و CD45 به ترتیب 8/2% و 8/3% بود. پس از تحریک MSCs با دگزامتازون، گلیسرول فسفات، آسکوربیک اسید و 10% FBS ، پتانسیل تمایزی این سلولها به استئوبلاستها ارزیابی شد. MSCs در روز 17 کشت، تمایز استئوژنیک را با افزایش بیان آلکالن فسفاتاز و تشکیل ماتریکس کلسیم خارج سلولی نشان دادند.
سنجش MTT : این سنجش نشان داد که µM 10 TEPA هیچ گونه اثر مشخصی بر روی تکثیر سلولی و حیات MSCs ندارد(نمودار1).
اثر TEPA بر میزان مس: غلظت مس سلولی پیش از افزودن TEPA در اندازهگیری با اتمیک ابزربشن ng 7 در 107 سلول بود. هنگامی که TEPA به محیط کشت اضافه شد، میزان مس سلولی به 4 نانو گرم در 107 سلول کاهش یافت. این نتایج نشان میدهند که میزان مس سلولی را میتوان با افزودن TEPA به محیط کشت کاهش داد.
ارزیابی تکثیر در شرایط کشت مختلف: بـه منظـور تعییـن اثـرات MSCs و کاهـش مس جهت حمایت از تکثیر سلولهای بنیادی خونساز، چهار وضعیت زیر مطرح بود: شرایط کشت اول؛ میانگین میزان چند برابر شدن TNCs، سلولهـای CD34+ و CFU-C در ایـن شرایــط به ترتیب 7/8 ± 48 ، 6/5 ± 7/18 و 7/12 ± 5/45 بود.
نمودار 1: اثر غظتهای مختلف TEPA بر توکسیسیتی و حیات سلولهای بنیادی مزانشیمی با استفاده از سنجش MTT ، رشد MSCs در حضور µM 10 TEPA به صورت معناداری بیشتر از سایر غلظتها بود(05/0 p<*).
نمودار 2: میانگین میزان چند برابر شدن پس از 10 روز تکثیر در شرایط کشت مختلف. A 2 : میانگین میزان چند برابر شدن TNCs ، B 2 : میانگین میزان چند برابر شدن سلولهای CD34+، C 2 : میانگین میزان چند برابر شدن سلولهای CD34+CD38-D 2 : میانگین میزان چند برابر شدن CFCs . شرایط کشت اول(1)، تنها در بردارنده سایتوکاینها بود. شرایط کشت دوم(2)، در بردارنده MSCs به عنوان لایه فیدر و سایتوکاینها بود. شرایط کشت سوم(3)، دربردارنده سایتوکاینها و TEPA بود. شرایط کشت چهارم(4)، در بردارنده MSCs به عنوان لایه فیدر، سایتوکاینها و TEPA بود. تفاوتهای معناداری بین شرایط کشت مختلف مشاهده گردید(05/0 p<*).
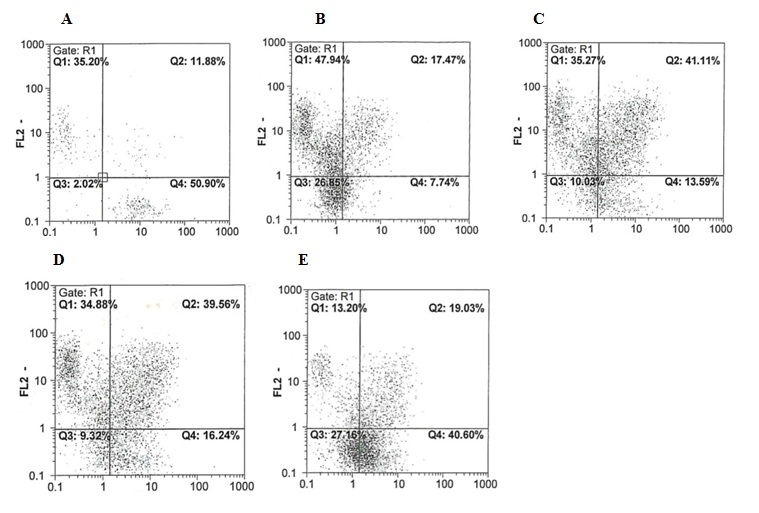
شکل 2 : آنالیز بیان CD34 و CD38 در سلولهای بنیادی خونساز قبل و پس از 10 روز تکثیر در شرایط کشت مختلف. FL1 : بیانگرCD34 ، FL2 : بیانگر CD38 . A2 : قبل از کشت(زمان صفر) B2 : شرایط کشت اول. C2 : شرایط کشت دوم. D2 : شرایط کشت سوم E2: شرایط کشت چهارم.
شرایط کشت دوم؛ در این شرایط اثر MSCs بر روی تکثیر TNCs ، سلولهـای CD34+ و CFU-C در سینــرژی بـا سایتوکاینها مورد مطالعه قرار گرفت. میانگین میزان چند برابر شدن آنها به ترتیب 2/15 ± 62 ، 4/11 ± 5/58 و 6/18 ± 6/78 بود. شرایط کشت سوم: به منظور ارزیابی اثر کاهش مس بر میزان تکثیر سلولی، سلولها با TEPA تیمار شدند. میانگین میزان چند برابر شدن TNCs ، سلولهای CD34+ و CFU-C به ترتیب 8/8 ± 52 ، 7/8 ± 45/44 و 4/15 ± 94/58 بود. شرایط کشت چهارم: اثر همکشتی MSCs در سینرژی با سایتوکاینها و TEPA بر روی تکثیر HSCs مورد ارزیابی قرار گرفت. پس از 10 روز کشت، تعداد TNCs ، سلولهای CD34+ و CFU-C در این شرایط به طور قابل توجهی افزایش یافت(05/0 p<). میانگین میزان چند برابر شدن TNCs، سلولهای CD34+ و CFU-C به ترتیب 87/21 ± 5/118 ، 3/15 ± 11/110 و 7/44 ± 9/172 بود. شرایط کشت مختلف به موازات هم با یکدیگر مقایسه شدند. تعداد CD34+ cells ، TNCs و CFU-C در روز دهم کشت مورد ارزیابی و مقایسه قرار گرفت(شکل 1 و نمودار 2).
آنالیز فلوسایتومتری پس از تکثیر: در این مطالعه بیان مارکرهای CD34 و CD38 توسط فلوسایتومتری ارزیابی شد. در شرایط کشت چهارم، سلولهای بیشتری با فنوتیپ ابتداییتر(05/0 p< ، 86/4% ± 7/39%) CD34+CD38- در مقایسـه بـا سه شرایط کشـت دیگر یافت شد(شکل 2). بحث تکثیر HSCs در ex vivo راهکار امید بخشی جهت اهداف تحقیقاتی و درمانی میباشد(36-34). در مطالعههای گذشته تلاش شده است تا تکثیر سلولهای CD34+ در شرایط کشت، بهینه شود(37، 17). HSCs را میتوان در محیطهای کشت همراه با سایتوکاینها تکثیر نمود اما این نوع از تکثیر همراه با تمایز میباشد(14). در مطالعههای مختلف نشان داده شده است که لایه فیدر استرومال قادر است از طریق ترشح فاکتورهای رشد و cell-to-cell contact ، از تکثیر HSCs حمایت کند (39، 38). از سوی دیگر در برخی مطالعهها نیز نقش TEPA در افزایش تکثیر و کاهش تمایز سلولهای بنیادی هماتوپوئتیک بند ناف بررسی شده است(25-22). به همین دلیل در این مطالعه بر آن شدیم تا اثرات همزمان MSCs به عنوان لایه فیدر و شلاتور مس جهت کاهش غلظت یون مس را بر میزان تکثیر سلولهای بنیادی خونساز بند ناف بررسی کنیم. به دنبال کشت سلولهای CD34+ در حضور سایتوکاینها به تنهایی(شرایط کشت اول)، میانگین میزان چند برابر شدن CD34+cells ، TNCs و CFU-C افزایش یافت(به ترتیب درحدود 48، 7/18 و 5/45 برابر) اما این افزایش در مقایسه با سایر شرایط کشت قابل توجه نبود. این نتایج با نتایج مطالعههای قبلی مطابقت داشت(42-40). به منظور تایید نقش MSCs بر روی تکثیر HSCs خون بند ناف، سلولهای CD34+ جدا شده، بر روی لایه فیدر سلولهای بنیادی مزانشیمی در حضور سایتوکاینها کشت داده شدند(شرایط کشت دوم). میزان تکثیر TNCs,CD34+cells و CFU-C در این شرایط بیشتر از تکثیر در شرایط کشت اول بود. مقایسه نتایج این دو شرایط کشت نشان میدهد که در طول کشت، MSCs باعث افزایش تکثیر TNCs و حفظ HSCs در حالت ابتدایی میشوند. مطالعهها نشان دادهاند که MSCs انواعی از سیگنالها(لیگاندهای چسبندگی، سایتوکاینها و پروتئینهای ماتریکس خارج سلولی)، فاکتورهای رونویسی و مولکولهای سیگنالدهی را جهت تنظیم تکثیر، تمایز و بقای HSCs در بدن و در شرایط آزمایشگاهی فراهـم میکنند(43). بـه نظـر مـیرسد تمـاس مستقیم بین HSCs و MSCs ، امری مهم برای حفظ stemness بوده و تاثیری قابل توجه بر روی عملکرد، فنوتیپ، مهاجرت و پروفایل بیان ژنی HSCs در شرایط آزمایشگاهی دارد(46-44، 19). آلاکل و همکاران نشان دادند که Non adherent HSCs از نظر عملکرد، فنوتیپ و کلونیزایی با adherent HSCs متفاوت میباشند. آنها نشان دادند که فراکسیون سلولی چسبنده متشکل از سلولهای ابتداییتر میباشد، از این رو MSCs دارای اثرات قابل توجهی بر روی HSCs در سیستمهای آزمایشگاهی میباشند. HSCs ابتداییتر دارای تمایل بیشتری به MSCs میباشند(47). در شرایط کشت اول، بسیاری از سلولهای اولیه تمایز یافته و مارکر سطحی CD34 خود را از دست میدهند. از این رو MSCs جهت تکثیر HSCs ضروری میباشند. مس در بسیاری از فرآیندهای سلولی از جمله تنظیم بیان ژنها، عملکرد پروتئینها و تکثیر و تمایز سلولی دخیل میباشد. با این حال به دلیل نقش آن در تولید گونههای فعال اکسیژن(ROSs) و افزایش مس سلولی، توکسیک میباشد(48). در شرایطی که سلولهای CD34+ در حضور سایتوکاینها و TEPA (شرایط کشت سوم) کشت داده شده بودند افزایش میزان تکثیر TNCs ، CD34+cells و CFU-C مشاهده شد(به ترتیب 52 ، 45/44 و 94/58 برابر). هر چند میزان تکثیر در این شرایط بیشتر از شرایط کشت اول بود اما کمتر از تکثیر در شرایط دوم بود. احتمالاً کاهش تکثیر ناشی از غیابcell-to-cell contact و سایتوکاینهای مترشحه از MSCs میباشد. مشخص شده است که فرآیندهای تکثیر و تمایز(به صورت متقابل) هر دو تحت تاثیر میزان مس قرار میگیرند اما در مقایسه با تکثیر، تمایز در اولویت قرار دارد(23). پلد و همکاران اثراتµM 10 TEPA را بر روی تکثیر سلولهای CD34+ ، مورد بررسی قرار دادند(25-23). در اولین مطالعه در سال 2001، این گروه اثرات TEPA، مس و سرولوپلاسمین را بر روی میزان مس سلولی و تکثیر سلولهای CD34+ ارزیابی کردند.در کشتهایی که در حضور سایتوکاینها و TEPA انجام شده بودند، تعداد سلولهای CD34+ ، TNCs و CFU-C بیشتر و میزان مس سلولی کمتر بود و در مقایسه با کشتهایی که تنها در حضور همان سایتوکاینها انجام شده بودند با تاخیر در تمایز همراه بودند، در حالی که کشتهای تیمار شده با مس نتایج بالعکسی را نشان دادند(23). نتایج ما با این نتایج نیز مطابقت داشت. پلد و همکاران در مطالعهای دیگر در سال 2004 اثر TEPA بر تکثیر سلولهای CD34+ و همچنین پتانسیل پیوند آنها به موشهای SCID را بررسی نمودند. نتایج آنها نشان داد که TEPA موجب افزایش تکثیر سلولها گردیده و از این رو توانایی پیوند را تقویت میکند(24). هر چند مکانیسم عملکرد TEPA نامشخص است اما نشان داده شده است که TEPA میزان رادیکالهای آزاد اکسیژن(ROS) و فعالیت سیتوکروم C اکسیداز(CCO) که برای حفظ HSCs در مغز استخوان ضروری هستند، را کاهش میدهد(50، 49). به علاوه در این مطالعه برای اولین بار سلولهای CD34+ در همکشتی با BM-MSCs به عنوان لایه فیدر و در حضور سایتوکاینها و TEPA (شرایط کشت چهارم) کشت داده شدند. میانگین میزان چند برابر شدن TNCs,CD34+cells و CFU-C در این شرایط به ترتیب 5/118، 11/110 و 9/172 بود. آنالیز ایمونوفنوتیپی HSCs در شرایط کشت مختلف نشان داد که در شرایط کشت چهارم، سلولهای CD34+CD38− تکثیر بهتر و تمایز کمتری را نسبت به سایر شرایط کشت دارا میباشند. در واقع این مطالعه نشان داد که کاهش میزان مس درون سلولی با استفاده از TEPA در همکشتی قادر است سلولهای CD34+ را با کارایی بهتری در مقایسه با سایر شرایط تکثیر نماید. کاربرد ترکیبی MSCs و TEPA میتواند راهکار جدیدی جهت افزایش تکثیر HSCs باشد. امید به این که با تحقیقات بیشتر، این استراتژی برای تکثیر HSCs و پیوند بهتر مفید واقع شود.
نتیجهگیری با توجه به نتایج به دست آمده از این مطالعه، اثر هم زمان سلولهای بنیادی مزانشیمی به عنوان لایه فیدر و کاهش یون مس جهت افزایش تعداد سلولهای بنیادی خونساز و حفظ پتانسیل کلونیزایی آنها در شرایط آزمایشگاهی مؤثر میباشد.
تشکر و قدردانی این تحقیق حاصل پایان نامه کارشناسی ارشد هماتولوژی است و در مرکز تحقیقات مؤسسه عالی آموزشی و پژوهشی طب انتقال خون تهران و با حمایت مالی دانشگاه علوم پزشکی تهران و مؤسسه عالی آموزشی و پژوهشی طب انتقال خون تهران به انجام رسیده است.
Amirizadeh N, Zaker F, Nasiri N. Evaluation of Umbilical Cord Blood CD34+ Hematopoietic Stem Cell Expansion in Co-culture with BM Mesenchymal Stem Cells in Presence of TEPA. Sci J Iran Blood Transfus Organ 2014; 10 (4) :353-363 URL: http://bloodjournal.ir/article-1-827-fa.html
امیریزاده ناصر، ذاکر فرهاد، نصیری ناهید. تکثیر سلولهای بنیادی خونساز CD34+ بند ناف در همکشتی با سلولهای بنیادی مزانشیمی مغز استخوان در حضور TEPA. فصلنامه پژوهشی خون. 1392; 10 (4) :353-363