

جلد 15، شماره 1 - ( بهار 1397 )
جلد 15 شماره 1 صفحات 70-59 |
برگشت به فهرست نسخه ها

![]()
![]()
![]()
Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Roozbahani M, Jamshidian H, Mahmoudi E, Arshi A. Angiogenesis: A Review of Molecular Mechanism. bloodj 2018; 15 (1) :59-70
URL: http://bloodjournal.ir/article-1-1141-fa.html
URL: http://bloodjournal.ir/article-1-1141-fa.html
روزبهانی ملیحه، جمشیدیان حسن، محمودی اسماعیل، عرشی اصغر. رگزایی، مروری بر مکانیسم مولکولی. فصلنامه پژوهشی خون. 1397; 15 (1) :59-70


باشگاه پژوهشگران جوان و نخبگان ـ واحد نجف آباد دانشگاه آزاد اسلامی
متن کامل [PDF 645 kb]
(13562 دریافت)
| چکیده (HTML) (10913 مشاهده)
مقدمه
واژه رگزایی به معنی ایجاد مویرگهای جدید از عروق موجود است. میتوان رگزایی را یک فرآیند ضروری در فیزیولوژی بدن دانست که با واسطه تعادل بین فاکتورهای القاکننده و مهارکننده رگزایی تنظیم میگردد و در صورتی که این تعادل از بین برود، زمینه برای بروز برخی بیماریها از جمله رشد و متاستاز تومور فراهم میشود(1). این فرآیند نیازمند تکثیر فعال سلولهای اندوتلیال است. به این صورت که تشکیل رگهای فعال، مستلزم برهمکنشهای هماهنگ بین سلولهای اندوتلیال، ماتریکس خارج سلولی و سلولهای احاطهکننده آنها میباشد(2).
مهمترین محرکهای فیزیولوژیکی رگزایی ایسکمی بافتی، هیپوکسی و التهاب هستند و علاوه بر آن برخی از فاکتورهای اختصاصی از قبیل فاکتور رشد رگی، سایتوکاینهـای التهابـی، مـولکولهای چسباننـده و نیتریـک اکساید، رگزایـی را تحریک و یا مهار میکنند(3).
متاستاز نقش ویژهای در گسترش سرطانهایی دارد که منجر به مرگ میگردند، در طی روند متاستاز سلولهای سرطانی از طریق عروق خونی مهاجرت نموده و به سایر بافتها وارد میشوند و در نهایت باعت درگیر شدن بافتهای سالم بدن میگردند(2، 1).
در سال 1971 فولکمن خاطر نشانکرد که "تومورها هرگز فراتر از اندازه مشخصی رشد نمیکنند مگر این که عروق آنها افزایش یابد". در این مقاله، او همچنین نظریهای را مطرح کرد مبنی بر این که تومورها دارای رگهای خونی جدیدی هستند که فاکتوری قابل انتشار را به نوعی به کار میگیرند. او از این فاکتورها به عنوان فاکتور آنژیوژنزی تومور یاد میکند.
در نهایت، اظهار داشت که از لحاظ تئوری، اگر بتوان رگزایی را مهار نمود، تومورها در اندازه کوچک باقی میمانند و سرانجام آسیبرسان نخواهند شد. از این رو مطابق با نظریه فولکمن، مهار رگزایی و متعاقب آن مهار متاستاز سلولها روش مناسبی برای مقابله با سرطان است و شناخت عوامل درگیر در رگزایی طبیعی و یا غیر طبیعی بسیار مهم و حیاتی میباشد(4).
روند رگزایی:
1- واسکولوژنز:
در ابتداییترین مراحل جنینی، جنین در فقدان عروق، توسعه و مواد غذایی را از طریق انتشار دریافت میکند. سپس در یک روند منظم و پیدرپی، جنین به سرعت تبدیل به موجودی با عروق بسیار میشود. سازماندهی اولیه سلولهای اندوتلیال که منجر به ایجاد عروق میگردد واسکولوژنژ خوانده شده و قبل از آن هیچ سیستم عروقی دیگری وجود ندارد(5).
لذا زمانی که بافت جدیدی تشکیل میشود، عروق خونی نیز بایستی توأم با آن به وجود آیند. واسکولوژنز با تمایز سلولهای مزودرمی به همانژیوبلاستها (که پیشساز سلولهای هماتوپوئتیک و سلولهای اندوتلیال هستند) آغاز میگردد(6).
با تمایز بیشتر، همانژیوبلاستها به آنژیوبلاست تبدیل شده و با تجمع آنژیوبلاستها جزایر خونی اولیه تشکیل میگردد. سپس این جزایر خونی با هم ادغام شده و شبکه اولیه عروقی که شامل مویرگهای نازک تشکیل شده توسط سلولهای اندوتلیال است پدیدار میشود. واسکولوژنز با تشکیل شبکه عروقی اولیه و تغییر شکل آن در حین فرآیند رگزایی ادامه مییابد(7، 6).
2- رگزایی:
رگزایی به ایجاد عروق جدید از عروق موجود گفته میشود که به دو شکل جوانهزدن و غیر جوانهزدن اتفاق میافتد. روش جوانهزنی به عنوان مکانیسم اصلی رگزایی در طی مراحل تکوین طبیعی و سرطانها محسوب میگردد(8). جوانه زدن به شاخهدار شدن و بیرونزدگی یک مویرگ جدید از مویرگ قبلی اشاره دارد که تکثیر بیش از حد سلولهای آندوتلیال، لازمه این رخداد است(9). برعکس روش غیرجوانهزدن، به دونیمه شدن رگ تکامل یافته با شکافت مویرگ از داخل(تقسیم طولی مویرگ) و تبدیل یک مویرگ به دو مویرگ اشاره دارد. در این روش، چون نیاز کمتری به تکثیر سلول آندوتلیال است در مقایسه با جوانه زدن فرآیندی کاراتر میباشد(11، 10).
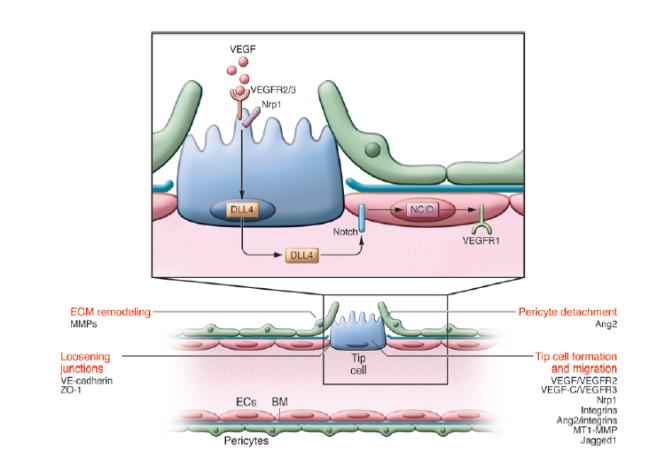
شکل 1: جوانه زنی رگ، انتخاب سلول رأس(10). سلولهای اندوتلیال با سیگنال های VEGFR2فعال شده، بیان Dll4را افزایش میدهند و به رسپتورNotch که روی سلولهای اندوتلیال همسایه است متصل میشود. سلولهای اندوتلیال با بالاترین VEGFR2 و کمترینVEGFR1 به موقعیت سلول رأس مهاجرت میکنند.
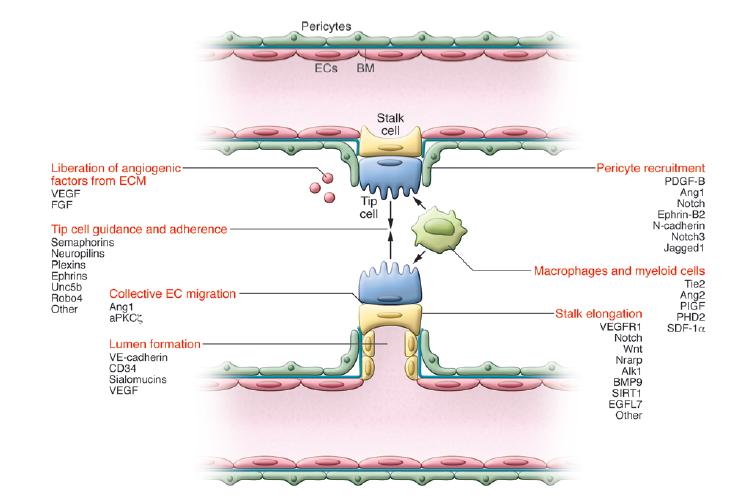
شکل 2: راهنمایی سلول رأس و طویل شدن ساقه(10). هنگامی که جوانههای در حال رشد در امتداد شیب VEGF حرکت میکنند، سلولهای نوک از طریق اینتگرین به ECM میچسبند و به سمت مولکولهای سیگنال هدایتگر مانند سمافورین و افرینها مهاجرت میکنند. سلولهای ساقه پشت سلول رأسی قرار میگیرند و برای طویل شدن جوانه و تشکیل لومن تکثیر میشوند.
3- آرتریوژنز:
فرآیند دیگری نیز در خصوص رگزایی به نام آرتریوژنز وجود دارد. آرتریوژنز به معنی بزرگ شدگی رگها هم از لحاظ قطر و هم از لحاظ ضخامت دیواره عروقی است (13، 12). آرتریوژنز عمدتاً در آرتریولهای بزرگ و آرتریتهای کوچک صورت میگیرد. آرتریوژنز مستلزم تکثیر سلولهای عضله صاف و سلولهای آندوتلیال است. مهمترین محرکهای درگیر در فرآیند آرتریوژنز، محرک همودینامیکی شیر استرس (فشار تنشی) است(15، 14).
مکانیسم مولکولی رگزایی:
نگاه کلی به مکانیسم مولکولی رگزایی:
محققان بر این عقیدهاند که برای القای رگزایی در شرایط فیزیولوژیک یا پاتولوژیک که مشتمل بر مراحل متعددی است، کاهش فشار اکسیژن(هیپوکسی) در بافت از اهمیت زیادی برخوردار است. در چنین شرایطی بافت دچار هیپوکسی، اقدام به ساخت و رهاسازی فاکتورهای رگزا همچون فاکتور رشد سلولهای اندوتلیال(VEGF) میکند(16). این فاکتورها پس از اتصال به گیرندههای خود بر روی سلولهای اندوتلیال، منجر به فعال شدن آنها میشوند. شاخص فعال شدن سلولهای اندوتلیال با شاخص میتوزی بالا، افزایش ظرفیت برای تهاجم و پروتئولیز ماتریکس شناخته میشود(17). سلولهای اندوتلیال فعال قادر به تهاجم به غشای پایه(BM) و ماتریکس خارج سلولی(ECM) و اختلال در اتصالات محکم، اتصالات چسبنده و gap junction در بین EC ها میباشد. به این صورت که با شروع فعالیت سلولهای اندوتلیال، انواع خاصی از متالوپروتئازها از سلولهای فوق ترشح میشود و غشای پایه را در منطقه مذکور تجزیه میکند(18). با هضم غشای پایه، سلولهای اندوتلیال اقدام به مهاجرت و تکثیر مینمایند، علاوه بر این، مولکولهای اتصالی از قبیل اینتگرین αvβ5 وαvβ3 نیز به فرآیند کشیدن و جلو رفتن جوانههای رگهای خونی در حال رشد کمک میکنند(19). در مراحل بعدی فرآیند رگزایی، متالوپروتئینازهای ماتریکس(Matrix Metalloproteinases; MMP) جهت تجزیه ماتریکس خارج سلولی و آغاز بازسازی مجدد آن تولید میشوند. سپس با برهم کنش آنژیوپوئیتین-Tie2 فرآیند تشکیل لوله آغاز میگردد. در مرحله بعد، سیستمEphB-ephrinB نیز تنظیم فرآیند تشکیل لولهها را بر عهده گرفته و در نهایت پـریسیتها و
سلولهای ماهیچهای صاف برای پایدار کردن رگخونی تازه
تشکیل شده، به این ساختار اضافه میشوند(21، 20).
مراحل مکانیسم مولکولی رگزایی:
1- جوانهزنی رگ:
1-1- انتخاب و هدایت سلول رأسی:
جوانه زنی رگ نیازمند همکاری بین مهاجرت سلولهای رأسی و تکثیر سلولهای ساقه است(شکل 1). در ابتدا سلولهای اندوتلیال موجود در لبه پیشرو جوانه رگی، میلههای میکرونی خود موسوم به فیلوپدیا را گسترش داده و به سمت سیگنالهای محرک رگزایی مهاجرت میکنند(22). به این صورت که در قسمت جلو که سطح VEGF دارای بالاترین مقدار است VEGF، رسپتور VEGFR2 موجود در سطح سلول رأس را برای تحریک مهاجرت سلول رأس فعال میکند. علاوه بر این سیگنالهای VEGFR2 از طریق یک کورسپتور به نام Nrp1 افزایش مییابد و باعث ارتقای عملکرد سلول رأسی میشود(23). اما چیزی که در این جا از اهمیت بالایی برخوردار است این است که اگر تمام سلولهای اندوتلیال به یک اندازه به محرکهای رگزایی واکنش نشان دهند، پس از آن قسمتی از شبکه عروقی از هم میپاشد و فراهم کردن خون در آن قسمت از بافت مختل میشود(24). در نتیجه برای جلوگیری از این اتفاق، مکانیسمهایی برای انتخاب تنها یک سلول اندوتلیال داخل مویرگ برای آغاز گسترش رگزایی(سلول رأس)، ایجاد شده است(25). از این مکانیسمها میتوان به رسپتورهای خانوادهNotch و لیگاندهایشان(Dll4) اشاره کرد. سلولهای اندوتلیال یا به سلولهای رأسی پیشرو و مهاجم تبدیل میشوند و یا به سلولهای تکثیرشونده ساقه، که این فنوتیپ را Notch تنظیم میکند(26). به این صورت که سلولهای اندوتلیال با سیگنالهای VEGFR2فعال شده، بیان Dll4را افزایش میدهند و به رسپتورNotch که روی سلولهای اندوتلیال همسایه است متصل میشوند. بعد از اتصالDll4 به گیرنده Notch روی سلولهای اندوتلیال همسایه، یک فاکتور رونویسی به نام NICD آزاد میشود که این فاکتور بیانVEGFR2 وNrp1 را کاهش میدهد در حالی که بیان VEGFR1 را افزایش میدهد. VEGFR1 با به دام انداختن VEGF، عدم پاسخ سلولهای ساقه را به VEGF کاهش میدهد. VEGFR1 درحالیکه تمایلش برای گرفتن VEGF از VEGFR2 بیشتر است اما قدرت سیگنالدهی آن ده برابر ضعیفتر از VEGFR2است. از این رو سلولهای اندوتلیال با بالاترین VEGFR2 و کمترینVEGFR1 به موقعیت سلول رأس مهاجرت میکنند(28، 27).
شرایط لازم برای مهاجرت سلول رأس:
تغییر وضعیت ECM
شل شدن اتصالات
کناره گیری پرسیتها
در نهایت تشکیل سلول رأس و مهاجرت(29)
2-1- تکثیر سلولهای ساقه و تشکیل لومن:
هنگامی که جوانههای در حال رشد در امتداد شیب VEGF حرکت میکنند، سلولهای نوک از طریق اینتگرین به ECM میچسبند و به سمت مولکولهای سیگنال هدایتگر مانند سمافورین و افرینها مهاجرت میکنند. سلولهای ساقه پشت سلول رأسی قرار میگیرند و برای طویل شدن جوانه و تشکیل لومن تکثیر میشوند(30). در حالی که سیگنالهای Notch مانع از تکثیر سلولهای ساقه میشود، بیان Nrarp در محل شاخهشکن به سیگنالهایWnt اجازه پشتیبانی از تکثیر سلولهای ساقه را میدهد. تشکیل لومن و متعاقب آن تشکیل لوله یکی از مشخصههای رگزایی و یک رفتار نسبتا خاص از سلولهای اندوتلیوم و اپیتلیوم است. با تکثیر سلولهای ساقه، لومن تا جایی طویل میشود که دو سلول رأسی از دو رگ مجاور به هم میرسند و ادغام میشوند. مکانیسم ادغام شدن توسط ماکروفاژهایی که در اطراف این رگها حضور دارند، انجام میشود(31). ماکروفاژها به عنوان سلولهای پل و از طریق تعامل داشتن با فیلوپدیاهای سلولهای رأسی مجاورشان، کار خود را انجام میدهند. اتصال و ارتباط سلولهای رأسی به همدیگر، توسط اتصالات حاوی VE-cadherin قویتر میشود و مـاکروفاژهای اطـراف عـروق از طریـق تولیـد فاکتورهای

شکل 3: بلوغ با واسطه پایداری سلولهای فالانکس(10).
سلولهای اندوتلیال از طریق سیگنالهای پاراکرین و اتوکرین و سلولهای حمایتکننده اطراف از طریق Notch ، Ang1 ، FGF و VEGF از فنوتیپ سلولهای اندوتلیال پایدار نگهداری و از رگها در مقابل شیب استرس حفاظت میکند.

شکل 4: درمانهای ضد رگزایی علیه VEGF و رسپتورهایش(13).
تعدادی از عوامل مهارکننده:
تا به امروز تعداد زیادی از فاکتورهای درونزای (Endogenous) مهارکننده رگزایی شناسایی شدهاند که منشاء بسیاری از آنها به طور طبیعی از ماتریکس خارجسلولی میباشد و برخی هم درواقع پروتیینهای غشای پایه هستند. بر این اساس، مهارکنندههای درونزای رگزایی به دو کلاس اصلی شامل مهارکنندههای مشتق از ماتریکس و مهارکنندههای مشتق نشده از ماتریکس تقسیم میشوند(51).
الف ـ مهارکنندههای مشتق از ماتریکس: 1- کانستاتین 2-
ترومبوسپوندین 1 و 2، 3- اندوستاتین 4- اجزای کلاژن و اجزای فیبرونکتین 5- اندورپلین 6- آرستن(51).
ب- مهارکنندههای مشتقنشده از ماتریکس: 1- آنژیواستاتین 2- آنتیترومبین 3- TIMP 4- اینترفرون 5- اینترلوکین 6- وازوستاتین 7- تروپونین 1، 8- اجزای پرولاکتین 9- SFlt-1 (51).
مکانیسم تاثیر مهارکنندههای رگزایی:
مهارکنندههای رگزایی از چند طریق پدیده رگزایی را مورد هدف قرار میدهند:
1- مهار مسیر VEGFشامل الف) مهارکنندههایPDGF وVEGF که گیرندههای تیروزین کیناز مانند Sunitinib، Sorafenib ،PD173074 ،SU6668 ، SU5416 و FGF را مهار میکنند ب) آنتیبادیهای مونوکلونال مهارکننده VEGF و گیرنده آن.
2- مهارکنندههـای مـاتریکـس متـالوپروتئینازهـا مانندBAY ،AG3340 ، .Marimastat
3- مهارکنندههای درونزاد مانند اندواستاتین، آنژیواستاتین،
تومستاتین، ترومبواسپوندین.
4- مـهـارکـننـدههـای کـیـنازهـای درون سلـولـی ماننــد
مهارکنندههای mTOR، مهارکنندههای پروتئین کینازC.
5- آنتاگونیستهای اینتگرین مانند Vitaxin، آنتی اینتگرین آنتیبادی، پپتیدهای مسدود کننده عملکرد α5β1 و ανβ3 اینتگرین یا Cilengitide.
6- سایتوکاینها و کموکاینها مانند فاکتور نکروز تومور، اینترفرون آلفا و بتا و اینترلوکین(52).
در زیر به برخی از این مکانیسمها اشاره شده است(22، 10):
1- مرگ برنامهریزی شده سلول
2- رونویسی فاکتورهای رشد در سلولهای سرطانی
3- انتقال سیگنال در سلولهای اندوتلیال
4- مهاجرت سلولهای اندوتلیال
5- بیان مولکولهای چسبان به نامE-cadherin
بسیاری از ترکیبات ضد رگزایی که اکنون در مرحله آزمایشهای کلینیکی قرار دارند، ترکیبات طبیعی هستند. تولید دارو از فرآوردههای طبیعی به سرعت در حال رشد و توسعه میباشد، این ترکیبات استراتژی بسیار امیدبخش برای شناسایی عوامل ضدرگزایی و ضد سرطان میباشند(53)(شکل 4).
نتیجهگیری
تهاجم و متاستاز از مشخصههای بیولوژیک تومورهای بدخیم و علت عمده عوارض جسمی و مرگ و میر سرطان میباشد. ادامه رشد نئوپلاسم اولیه و متاستاز بستگی به خونرسانی کافی به آن منطقه دارد. فرآیند تشکیل عروق جدید یا همان رگزایی، به تومورها این امکان را میدهد که فراتر از 1-2 میلیمتر مکعب توسعه یابند. به استثنای تومورهای خوشخیم که رگزایی کمی دارند و سرعت رشد آنها کند است، رگزایی ﻳﻚ ﻓﺮآﻳﻨﺪ ﻣﻬﻢ در ﺷﺮاﻳﻂ ﻓﻴﺰﻳﻮﻟﻮژﻳﻜﻲ و پاتولوژیکی است. گسترش سیستم عروقی، احتمال تهاجم سلولهای توموری را از طریق وارد شدن به جریان خون و انتشار به اندامهای دیگر افزایش میدهد. علاوه بر این، نشان داده شده که تشکیل سیستم عروقی در تومورهای بدخیم با قدرت متاستاز تومور رابطه مستقیم دارد. به طور کلی در بافتهای سالم و پایدار (Quiescent) فاکتورهایی که از رگزایی ممانعت میکنند غالب هستند اما در بافتهایی که به سرعت تقسیم میشوند، مولکولهایی که فرآیند رگزایی را تحریک میکنند غلبه دارند. به ﻫﻤﯿﻦ دﻟﯿﻞ ﻣﻬﺎر رگزایی ﺑﻪ ﻋﻨﻮان یﮏ ﻋﺎﻣﻞ کمکﮐﻨﻨﺪه در درﻣﺎن سرطان شناخته میشود. در نتیجه مطالعههای محققان ﺟﻬﺖ تشخیص ﻣﻜﺎﻧﻴﺴﻢ مولکولی و ﻓﺎﻛﺘﻮرﻫﺎی دﺧـﻴﻞ در اﻳﻦ ﻓﺮآﻳﻨﺪ، ﻣﻲﺗﻮاﻧﺪ زﻣﻴﻨﻪﺳﺎز ﺗﻮﺳﻌﻪ راهﻫﺎی درمانی باشد. با توجه به اهمیت رگزایی در تحقیقات مربوط به کشف و شناسایی فاکتورهای آنژیوژنیک و عوامل مهارکننده رگزایی جهت درمان
بیماریهای مختلف از جمله انواعی از تومورها که با رگزایی ارتباط تنگاتنگی داشته و به آن وابسته هستند، روشهـای مهـار رگزایی که با هدف تداخل با این فرآیند مهم جهتگیری نمودهاند، مسیر امیدوار کنندهای برای درمان سرطان محسوب میگردند. از طرف دیگر، ضرورت مطالعههای بیشتر در زمینه فرآوردههای طبیعی ضد رگزایـی و هم چنیـن مسیرهای مولکولی که منجر به مهار
رگزایی میشود، کاملاً محسوس به نظر میرسد.
تشکر و قدردانی
محققان و نویسندگان این مقاله بدینوسیله مراتب امتنان
خود را از همه عزیزانی که ما را در به ثمر رساندن این مقاله یاری نمودند، اعلام میدارند.
متن کامل: (19377 مشاهده)
رگزایی، مروری بر مکانیسم مولکولی
ملیحه روزبهانی1، حسن جمشیدیان2، اسماعیل محمودی3، اصغر عرشی4
چکیده
سابقه و هدف
رگزایی به تشکیل عروق خونی جدید از عروق موجود اطلاق میشود. رگزایی یک رویداد مهم در انواع فرآیندهای فیزیولوژیکی مانند رشد جنین، چرخه ماهیانه، تولید مثل و ترمیم زخم است. هم چنین در تعدادی از فرآیندهای پاتولوژیکی مانند تومور، بیماری چشم، رتینوپاتی، نفروپاتی دیابتی و چاقی دارای نقش مرکزی است. رگزایی توسط ترکیبی از عوامل رشد و محرکهای رگزایی هدایت و توسط گروهی به همان اندازه متنوع از مهارکنندههای رگزایی، تعدیل میشود.
مواد و روشها
به دلیل اهمیت فرآیند رگزایی در ایجاد بیماریهای وابسته به آن، در این مقاله مروری به بررسی ابعاد مختلف فرآیند رگزایی و مکانیسمها و عوامل مربوط به آن و هم چنین مطالعههای پیرامون آنها که بالغ بر 50 مقاله میباشد، پرداخته شده است. برای جستجوی مقالهها از پایگاههای خارجی و داخلی مورد اطمینان نظیر NCBI و SID استفاده شد.
یافتهها
بر اساس کارهای اخیر از چندین آزمایشگاه، واضح است که فرآیندهای رگزایی و بیماریهای مرتبط با ایجاد رگ، نه تنها نتیجه تولید نامحدود فرمهای طبیعی و غیر طبیعی از واسطههای محرک رگزایی است بلکه نتیجه کمبود نسبی مولکولهای مهارکننده رگزایی نیز میباشد. به طور کلی، این فرآیند تحت تاثیر عوامل مختلف بوده و دربرگیرنده یک سری رخدادهای سلولی از قبیل مهاجرت، تکثیر و تمایز سلولهای اندوتلیال و در نهایت تشکیل عروق، بلوغ و بازسازی نهایی آنها است.
نتیجه گیری
فاکتور کلیدی و مؤثر در تکثیر و مهاجرت سلولهای اندوتلیال که اساس تشکیل هر رگ جدیدی میباشد، فاکتور رشد اندوتلیوم عروقی است. رگزایی درمانی شامل مهار رگزایی غیر طبیعی در مواردی مثل تومورها و تحریک رگزایی در بیماریهای ایسکمی مثل ایسکمیک قلبی و یا بیماریهای عروق محیطی است.
کلمات کلیدی: پاتولوژی، فاکتور رشد اندوتلیوم عروقی، رگهای خونی
تاریخ دریافت: 6 /6 /96
تاریخ پذیرش: 19/10/96
1- کارشناس ارشد ژنتیک ـ دانشکده علوم پایه دانشگاه آزاد اسلامی ـ واحد شهرکرد ـ شهرکرد ـ ایران
2- کارشناس ارشد ژنتیک ـ مرکز تحقیقات انتقال خون، مؤسسه عالی آموزشی و پژوهشی طب انتقال خون و پایگاه منطقهای انتقال خون اصفهان ـ اصفهان ـ ایران
3- کارشناس ارشد میکروبیولوژی ـ مرکز تحقیقات بیوتکنولوژی دانشگاه آزاد اسلامی واحد شهرکرد ـ شهرکرد ـ ایران
4- مؤلف مسئول: کارشناس ارشد ژنتیک ـ باشگاه پژوهشگران جوان و نخبگان ـ واحد نجفآباد دانشگاه آزاد اسلامی ـ نجفآباد ـ ایران ـ صندوق پستی: 517
ملیحه روزبهانی1، حسن جمشیدیان2، اسماعیل محمودی3، اصغر عرشی4
چکیده
سابقه و هدف
رگزایی به تشکیل عروق خونی جدید از عروق موجود اطلاق میشود. رگزایی یک رویداد مهم در انواع فرآیندهای فیزیولوژیکی مانند رشد جنین، چرخه ماهیانه، تولید مثل و ترمیم زخم است. هم چنین در تعدادی از فرآیندهای پاتولوژیکی مانند تومور، بیماری چشم، رتینوپاتی، نفروپاتی دیابتی و چاقی دارای نقش مرکزی است. رگزایی توسط ترکیبی از عوامل رشد و محرکهای رگزایی هدایت و توسط گروهی به همان اندازه متنوع از مهارکنندههای رگزایی، تعدیل میشود.
مواد و روشها
به دلیل اهمیت فرآیند رگزایی در ایجاد بیماریهای وابسته به آن، در این مقاله مروری به بررسی ابعاد مختلف فرآیند رگزایی و مکانیسمها و عوامل مربوط به آن و هم چنین مطالعههای پیرامون آنها که بالغ بر 50 مقاله میباشد، پرداخته شده است. برای جستجوی مقالهها از پایگاههای خارجی و داخلی مورد اطمینان نظیر NCBI و SID استفاده شد.
یافتهها
بر اساس کارهای اخیر از چندین آزمایشگاه، واضح است که فرآیندهای رگزایی و بیماریهای مرتبط با ایجاد رگ، نه تنها نتیجه تولید نامحدود فرمهای طبیعی و غیر طبیعی از واسطههای محرک رگزایی است بلکه نتیجه کمبود نسبی مولکولهای مهارکننده رگزایی نیز میباشد. به طور کلی، این فرآیند تحت تاثیر عوامل مختلف بوده و دربرگیرنده یک سری رخدادهای سلولی از قبیل مهاجرت، تکثیر و تمایز سلولهای اندوتلیال و در نهایت تشکیل عروق، بلوغ و بازسازی نهایی آنها است.
نتیجه گیری
فاکتور کلیدی و مؤثر در تکثیر و مهاجرت سلولهای اندوتلیال که اساس تشکیل هر رگ جدیدی میباشد، فاکتور رشد اندوتلیوم عروقی است. رگزایی درمانی شامل مهار رگزایی غیر طبیعی در مواردی مثل تومورها و تحریک رگزایی در بیماریهای ایسکمی مثل ایسکمیک قلبی و یا بیماریهای عروق محیطی است.
کلمات کلیدی: پاتولوژی، فاکتور رشد اندوتلیوم عروقی، رگهای خونی
تاریخ دریافت: 6 /6 /96
تاریخ پذیرش: 19/10/96
1- کارشناس ارشد ژنتیک ـ دانشکده علوم پایه دانشگاه آزاد اسلامی ـ واحد شهرکرد ـ شهرکرد ـ ایران
2- کارشناس ارشد ژنتیک ـ مرکز تحقیقات انتقال خون، مؤسسه عالی آموزشی و پژوهشی طب انتقال خون و پایگاه منطقهای انتقال خون اصفهان ـ اصفهان ـ ایران
3- کارشناس ارشد میکروبیولوژی ـ مرکز تحقیقات بیوتکنولوژی دانشگاه آزاد اسلامی واحد شهرکرد ـ شهرکرد ـ ایران
4- مؤلف مسئول: کارشناس ارشد ژنتیک ـ باشگاه پژوهشگران جوان و نخبگان ـ واحد نجفآباد دانشگاه آزاد اسلامی ـ نجفآباد ـ ایران ـ صندوق پستی: 517
مقدمه
واژه رگزایی به معنی ایجاد مویرگهای جدید از عروق موجود است. میتوان رگزایی را یک فرآیند ضروری در فیزیولوژی بدن دانست که با واسطه تعادل بین فاکتورهای القاکننده و مهارکننده رگزایی تنظیم میگردد و در صورتی که این تعادل از بین برود، زمینه برای بروز برخی بیماریها از جمله رشد و متاستاز تومور فراهم میشود(1). این فرآیند نیازمند تکثیر فعال سلولهای اندوتلیال است. به این صورت که تشکیل رگهای فعال، مستلزم برهمکنشهای هماهنگ بین سلولهای اندوتلیال، ماتریکس خارج سلولی و سلولهای احاطهکننده آنها میباشد(2).
مهمترین محرکهای فیزیولوژیکی رگزایی ایسکمی بافتی، هیپوکسی و التهاب هستند و علاوه بر آن برخی از فاکتورهای اختصاصی از قبیل فاکتور رشد رگی، سایتوکاینهـای التهابـی، مـولکولهای چسباننـده و نیتریـک اکساید، رگزایـی را تحریک و یا مهار میکنند(3).
متاستاز نقش ویژهای در گسترش سرطانهایی دارد که منجر به مرگ میگردند، در طی روند متاستاز سلولهای سرطانی از طریق عروق خونی مهاجرت نموده و به سایر بافتها وارد میشوند و در نهایت باعت درگیر شدن بافتهای سالم بدن میگردند(2، 1).
در سال 1971 فولکمن خاطر نشانکرد که "تومورها هرگز فراتر از اندازه مشخصی رشد نمیکنند مگر این که عروق آنها افزایش یابد". در این مقاله، او همچنین نظریهای را مطرح کرد مبنی بر این که تومورها دارای رگهای خونی جدیدی هستند که فاکتوری قابل انتشار را به نوعی به کار میگیرند. او از این فاکتورها به عنوان فاکتور آنژیوژنزی تومور یاد میکند.
در نهایت، اظهار داشت که از لحاظ تئوری، اگر بتوان رگزایی را مهار نمود، تومورها در اندازه کوچک باقی میمانند و سرانجام آسیبرسان نخواهند شد. از این رو مطابق با نظریه فولکمن، مهار رگزایی و متعاقب آن مهار متاستاز سلولها روش مناسبی برای مقابله با سرطان است و شناخت عوامل درگیر در رگزایی طبیعی و یا غیر طبیعی بسیار مهم و حیاتی میباشد(4).
روند رگزایی:
1- واسکولوژنز:
در ابتداییترین مراحل جنینی، جنین در فقدان عروق، توسعه و مواد غذایی را از طریق انتشار دریافت میکند. سپس در یک روند منظم و پیدرپی، جنین به سرعت تبدیل به موجودی با عروق بسیار میشود. سازماندهی اولیه سلولهای اندوتلیال که منجر به ایجاد عروق میگردد واسکولوژنژ خوانده شده و قبل از آن هیچ سیستم عروقی دیگری وجود ندارد(5).
لذا زمانی که بافت جدیدی تشکیل میشود، عروق خونی نیز بایستی توأم با آن به وجود آیند. واسکولوژنز با تمایز سلولهای مزودرمی به همانژیوبلاستها (که پیشساز سلولهای هماتوپوئتیک و سلولهای اندوتلیال هستند) آغاز میگردد(6).
با تمایز بیشتر، همانژیوبلاستها به آنژیوبلاست تبدیل شده و با تجمع آنژیوبلاستها جزایر خونی اولیه تشکیل میگردد. سپس این جزایر خونی با هم ادغام شده و شبکه اولیه عروقی که شامل مویرگهای نازک تشکیل شده توسط سلولهای اندوتلیال است پدیدار میشود. واسکولوژنز با تشکیل شبکه عروقی اولیه و تغییر شکل آن در حین فرآیند رگزایی ادامه مییابد(7، 6).
2- رگزایی:
رگزایی به ایجاد عروق جدید از عروق موجود گفته میشود که به دو شکل جوانهزدن و غیر جوانهزدن اتفاق میافتد. روش جوانهزنی به عنوان مکانیسم اصلی رگزایی در طی مراحل تکوین طبیعی و سرطانها محسوب میگردد(8). جوانه زدن به شاخهدار شدن و بیرونزدگی یک مویرگ جدید از مویرگ قبلی اشاره دارد که تکثیر بیش از حد سلولهای آندوتلیال، لازمه این رخداد است(9). برعکس روش غیرجوانهزدن، به دونیمه شدن رگ تکامل یافته با شکافت مویرگ از داخل(تقسیم طولی مویرگ) و تبدیل یک مویرگ به دو مویرگ اشاره دارد. در این روش، چون نیاز کمتری به تکثیر سلول آندوتلیال است در مقایسه با جوانه زدن فرآیندی کاراتر میباشد(11، 10).
شکل 1: جوانه زنی رگ، انتخاب سلول رأس(10). سلولهای اندوتلیال با سیگنال های VEGFR2فعال شده، بیان Dll4را افزایش میدهند و به رسپتورNotch که روی سلولهای اندوتلیال همسایه است متصل میشود. سلولهای اندوتلیال با بالاترین VEGFR2 و کمترینVEGFR1 به موقعیت سلول رأس مهاجرت میکنند.
شکل 2: راهنمایی سلول رأس و طویل شدن ساقه(10). هنگامی که جوانههای در حال رشد در امتداد شیب VEGF حرکت میکنند، سلولهای نوک از طریق اینتگرین به ECM میچسبند و به سمت مولکولهای سیگنال هدایتگر مانند سمافورین و افرینها مهاجرت میکنند. سلولهای ساقه پشت سلول رأسی قرار میگیرند و برای طویل شدن جوانه و تشکیل لومن تکثیر میشوند.
3- آرتریوژنز:
فرآیند دیگری نیز در خصوص رگزایی به نام آرتریوژنز وجود دارد. آرتریوژنز به معنی بزرگ شدگی رگها هم از لحاظ قطر و هم از لحاظ ضخامت دیواره عروقی است (13، 12). آرتریوژنز عمدتاً در آرتریولهای بزرگ و آرتریتهای کوچک صورت میگیرد. آرتریوژنز مستلزم تکثیر سلولهای عضله صاف و سلولهای آندوتلیال است. مهمترین محرکهای درگیر در فرآیند آرتریوژنز، محرک همودینامیکی شیر استرس (فشار تنشی) است(15، 14).
مکانیسم مولکولی رگزایی:
نگاه کلی به مکانیسم مولکولی رگزایی:
محققان بر این عقیدهاند که برای القای رگزایی در شرایط فیزیولوژیک یا پاتولوژیک که مشتمل بر مراحل متعددی است، کاهش فشار اکسیژن(هیپوکسی) در بافت از اهمیت زیادی برخوردار است. در چنین شرایطی بافت دچار هیپوکسی، اقدام به ساخت و رهاسازی فاکتورهای رگزا همچون فاکتور رشد سلولهای اندوتلیال(VEGF) میکند(16). این فاکتورها پس از اتصال به گیرندههای خود بر روی سلولهای اندوتلیال، منجر به فعال شدن آنها میشوند. شاخص فعال شدن سلولهای اندوتلیال با شاخص میتوزی بالا، افزایش ظرفیت برای تهاجم و پروتئولیز ماتریکس شناخته میشود(17). سلولهای اندوتلیال فعال قادر به تهاجم به غشای پایه(BM) و ماتریکس خارج سلولی(ECM) و اختلال در اتصالات محکم، اتصالات چسبنده و gap junction در بین EC ها میباشد. به این صورت که با شروع فعالیت سلولهای اندوتلیال، انواع خاصی از متالوپروتئازها از سلولهای فوق ترشح میشود و غشای پایه را در منطقه مذکور تجزیه میکند(18). با هضم غشای پایه، سلولهای اندوتلیال اقدام به مهاجرت و تکثیر مینمایند، علاوه بر این، مولکولهای اتصالی از قبیل اینتگرین αvβ5 وαvβ3 نیز به فرآیند کشیدن و جلو رفتن جوانههای رگهای خونی در حال رشد کمک میکنند(19). در مراحل بعدی فرآیند رگزایی، متالوپروتئینازهای ماتریکس(Matrix Metalloproteinases; MMP) جهت تجزیه ماتریکس خارج سلولی و آغاز بازسازی مجدد آن تولید میشوند. سپس با برهم کنش آنژیوپوئیتین-Tie2 فرآیند تشکیل لوله آغاز میگردد. در مرحله بعد، سیستمEphB-ephrinB نیز تنظیم فرآیند تشکیل لولهها را بر عهده گرفته و در نهایت پـریسیتها و
سلولهای ماهیچهای صاف برای پایدار کردن رگخونی تازه
تشکیل شده، به این ساختار اضافه میشوند(21، 20).
مراحل مکانیسم مولکولی رگزایی:
1- جوانهزنی رگ:
1-1- انتخاب و هدایت سلول رأسی:
جوانه زنی رگ نیازمند همکاری بین مهاجرت سلولهای رأسی و تکثیر سلولهای ساقه است(شکل 1). در ابتدا سلولهای اندوتلیال موجود در لبه پیشرو جوانه رگی، میلههای میکرونی خود موسوم به فیلوپدیا را گسترش داده و به سمت سیگنالهای محرک رگزایی مهاجرت میکنند(22). به این صورت که در قسمت جلو که سطح VEGF دارای بالاترین مقدار است VEGF، رسپتور VEGFR2 موجود در سطح سلول رأس را برای تحریک مهاجرت سلول رأس فعال میکند. علاوه بر این سیگنالهای VEGFR2 از طریق یک کورسپتور به نام Nrp1 افزایش مییابد و باعث ارتقای عملکرد سلول رأسی میشود(23). اما چیزی که در این جا از اهمیت بالایی برخوردار است این است که اگر تمام سلولهای اندوتلیال به یک اندازه به محرکهای رگزایی واکنش نشان دهند، پس از آن قسمتی از شبکه عروقی از هم میپاشد و فراهم کردن خون در آن قسمت از بافت مختل میشود(24). در نتیجه برای جلوگیری از این اتفاق، مکانیسمهایی برای انتخاب تنها یک سلول اندوتلیال داخل مویرگ برای آغاز گسترش رگزایی(سلول رأس)، ایجاد شده است(25). از این مکانیسمها میتوان به رسپتورهای خانوادهNotch و لیگاندهایشان(Dll4) اشاره کرد. سلولهای اندوتلیال یا به سلولهای رأسی پیشرو و مهاجم تبدیل میشوند و یا به سلولهای تکثیرشونده ساقه، که این فنوتیپ را Notch تنظیم میکند(26). به این صورت که سلولهای اندوتلیال با سیگنالهای VEGFR2فعال شده، بیان Dll4را افزایش میدهند و به رسپتورNotch که روی سلولهای اندوتلیال همسایه است متصل میشوند. بعد از اتصالDll4 به گیرنده Notch روی سلولهای اندوتلیال همسایه، یک فاکتور رونویسی به نام NICD آزاد میشود که این فاکتور بیانVEGFR2 وNrp1 را کاهش میدهد در حالی که بیان VEGFR1 را افزایش میدهد. VEGFR1 با به دام انداختن VEGF، عدم پاسخ سلولهای ساقه را به VEGF کاهش میدهد. VEGFR1 درحالیکه تمایلش برای گرفتن VEGF از VEGFR2 بیشتر است اما قدرت سیگنالدهی آن ده برابر ضعیفتر از VEGFR2است. از این رو سلولهای اندوتلیال با بالاترین VEGFR2 و کمترینVEGFR1 به موقعیت سلول رأس مهاجرت میکنند(28، 27).
شرایط لازم برای مهاجرت سلول رأس:
تغییر وضعیت ECM
شل شدن اتصالات
کناره گیری پرسیتها
در نهایت تشکیل سلول رأس و مهاجرت(29)
2-1- تکثیر سلولهای ساقه و تشکیل لومن:
هنگامی که جوانههای در حال رشد در امتداد شیب VEGF حرکت میکنند، سلولهای نوک از طریق اینتگرین به ECM میچسبند و به سمت مولکولهای سیگنال هدایتگر مانند سمافورین و افرینها مهاجرت میکنند. سلولهای ساقه پشت سلول رأسی قرار میگیرند و برای طویل شدن جوانه و تشکیل لومن تکثیر میشوند(30). در حالی که سیگنالهای Notch مانع از تکثیر سلولهای ساقه میشود، بیان Nrarp در محل شاخهشکن به سیگنالهایWnt اجازه پشتیبانی از تکثیر سلولهای ساقه را میدهد. تشکیل لومن و متعاقب آن تشکیل لوله یکی از مشخصههای رگزایی و یک رفتار نسبتا خاص از سلولهای اندوتلیوم و اپیتلیوم است. با تکثیر سلولهای ساقه، لومن تا جایی طویل میشود که دو سلول رأسی از دو رگ مجاور به هم میرسند و ادغام میشوند. مکانیسم ادغام شدن توسط ماکروفاژهایی که در اطراف این رگها حضور دارند، انجام میشود(31). ماکروفاژها به عنوان سلولهای پل و از طریق تعامل داشتن با فیلوپدیاهای سلولهای رأسی مجاورشان، کار خود را انجام میدهند. اتصال و ارتباط سلولهای رأسی به همدیگر، توسط اتصالات حاوی VE-cadherin قویتر میشود و مـاکروفاژهای اطـراف عـروق از طریـق تولیـد فاکتورهای
شکل 3: بلوغ با واسطه پایداری سلولهای فالانکس(10).
سلولهای اندوتلیال از طریق سیگنالهای پاراکرین و اتوکرین و سلولهای حمایتکننده اطراف از طریق Notch ، Ang1 ، FGF و VEGF از فنوتیپ سلولهای اندوتلیال پایدار نگهداری و از رگها در مقابل شیب استرس حفاظت میکند.
رگزایـی و یـا آزادسـازی آنهـا ازECM ، جوانــهزدن را
بیشتر تحریک میکنند. سلولهای ساقه همچنین با رسوب غشای پایه و جذب پرسیتها، موجب پایداری رگ درحال تشکیل میشوند. پیشسازهای پرسیت(سلولهای پیشساز مزانشیمی) از طریق بیان PDGF از سلولهای اندوتلیال جذب رگها میشوند و در پاسخ به TGFβ به پرسیتها تمایز مییابند. پرسیتها با کاهش مهاجرت، تکثیر سلولهای اندوتلیال و کاهش نشت عروق منجر به پایداری رگهای نو ظهور میشوند(32).
شرایط لازم و فاکتورهای دخیل در تشکیل لومن و تکثیر ساقه(شکل 2):
آزادسازی فاکتورهای محرک رگزایی از ECM
راهنمایی و چسبندگی سلول رأسی
مهاجرت سلولهای اندوتلیال به هم پیوسته
طویل شدن ساقه
تشکیل لومن
به کارگیری ماکروفاژها و سلولهای میلوئید
استخدام پرسیتها(33، 32)
2- بلوغ به واسطه پایداری سلولهای فالانکس:
هنگامی که ادغام صورت میگیرد، یک لومن متصل به هم برای عبور جریان خون در سرتاسر مویرگ جدید به وجود میآید. عبور جریان خون و دریافت اکسیژن و مواد غذایی توسط بافت هیپوکسی منجر به کاهش سطح سیگنالهای رگزایی، غیر فعال شدن سنسورهای اکسیژن سلولهای اندوتلیال و افزایش مولکولهای محرک پایداری منجر به پایداری سلولهای اندوتلیال میشود(34). سلولهای اندوتلیال دوباره فنوتیپ فالانکس پایدار را به صورت یک تک لایه محکم و مقاوم با یک سطح ساده برای انتقال جریان خون و تنظیم جریان خون به بافتها، از سر میگیرند.
سلولهای اندوتلیال از طریق سیگنالهای پاراکرین و اتوکرین و سلولهای حمایتکننده اطراف از طریق Notch، Ang1، FGF و VEGF از فنوتیپ سلولهای اندوتلیال پایدار نگهداری و از رگها در مقابل شیب استرس حفاظت میکنند. در نهایت با پایداری و بلوغ، رگ سدی را بین خون و بافت اطراف جهت کنترل تبادل مایعات و مواد تشکیل میدهد(35)(شکل 3).
شرایط لازم برای بلوغ رگها:
برقراری دوباره اتصالات سلول به سلول
رسوب غشا پایه
بلوغ پرسیتها
پایداری
ایجاد سلولهای فالانکس(36)
نقش رگزایی در تومور و متاستاز:
سلولهای تومور جمعیتی از سلولهای میزبان هستند که توانایی تنظیم تکثیر خود را از دست دادهاند و بنابراین به مقدار نامحدود تکثیر میشوند. بافت توموری میتواند مواد تغذیهای و اکسیژن کافی را از طریق انتشار ساده تا محدوده 1 تا 2 میلیمتر جذب نماید و از این نقطه به بعد نیازمند ایجاد رگهای تغذیهکننده جدید میباشند. در طی فرآیندی که بسیار به رگزایی نرمال شباهت دارد، یک تومور میتواند تشکیل رگهای جدید را از شبکه مویرگی موجود القا کند(38، 37).
تعدادی از عوامل فعالکننده:
1- VEGF: فاکتور رشد اندوتلیوم به عنوان فاکتور نفوذپذیری عروق نیز شناخته میشود. این فاکتور یک پروتئین همودایمر باند شده به هپارین است که دارای وزنی معادل 45 کیلو دالتون میباشد. VEGF دارای 7 ایزوفرم PIGF ، A ، B ، C ، D ، E و F که نتیجه اسپلایسینگ متفات از ژن VEFG میباشد، است.VEGF از طریق 1- تنظیم ساخت DNA باعث تکثیر سلولهای اندوتلیال 2- افزایش مؤلفههای آنتیآپوپتیک مانند BcL2 و A1 باعث بقا 3- تخریب غشا پایه باعث مهاجرت 4- فسفریله کردن اجزاء چسبنده بین سلولهای اندوتلیال و اتصالات محکم باعث نفوذپذیری سلولهای اندوتلیال میشود(39).
2- رسپتورهای VEGFR: VEGF دارای سه نوع گیرنده تیروزین کینازی به نامهای VEGFR3 (flt4)، VEGFR1 (flt1) و VEGFR2 (KDR/flk1) میباشد که این گیرندهها بر روی سلولهای اندوتلیال عروق وجود دارند.VEGFA بهVEGFR1 و VEGFR2 متصل میشود. VEGFB و PIGF تنها به VEGFR1 متصل میشود وVEGFD و VEGFC محکم به VEGFR3 متصل میشوند.VEGFA با افینیتی بالاتری به VEGFR1 نسبت به VEGFR2 متصل میشود در حالی که فعالیت تیروزین کینازی VEGFR1ده برابر ضعیفتر از VEGFR2است(40).
3- FGF : فاکتور رشد فیبروبلاست است که شامل 9 عضو مجزا میباشد. از مهمترین FGF ها میتوان بهbFGF و aFGF اشاره کرد که به رسپتورهای تیروزین کینازی FGFR1 و FGFR2 متصل میشود. این فاکتور قویترین فاکتور رگزایی و قابل اتصال به هپارین و هپاران سولفات است. این فاکتور هم به صورت ذخیره شده در غشای پایه به عنوان یک منبع ذخیره و هم به صورت افزایش بیان در طول رگزایی دیده میشود. این فاکتور به طور کلی در مهاجرت و تمایز سلولهای اندوتلیال و پروتئولیز خارجسلولی نقش دارد.FGF ها میتوژن اختصاصی سلولهای اندوتلیال بوده که توسط سلولهای توموری و ماکروفاژهای جذب شده ترشح میشوند(41).
4- HGF : فاکتور رشد هپاتوسیت، که در کاهش آپوپتوز سلولهای اندوتلیال و ترمیم ماتریکس خارجسلولی نقش دارد(15).
5- اینترلوکین 8: باعث تکثیر و مهاجرت سلولهای اندوتلیال و تشکیل تیوپ مویرگی میشود(16).
6-TNFα : یک سایتوکاین التهابی است که در ابتدا توسط ماکروفاژهای فعال شده در طول التهاب و پاسخهای ایمنی ترشح میشود. این فاکتور همچنین با مست سلها، سلولهای اندوتلیال، فیبروبلاستها و مونوسیت مرتبط است. TNFαبه طور مستقیم از طریق القای تمایز و به طور غیرمستقیم از طریق تحریک تولید فاکتورهای رگزایی از دیگر سلولها بر روی سلولهای اندوتلیال اثر میگذارد(42).
7- TGFβ: شامل سه ایزوتایپ TGFβ1، TGFβ2و TGFβ3 میباشد. این فاکتور در بدن باعث افزایش رسوب ECM و افزایش بیان رسپتورهای اینتگرین و واسطه تکثیر سلولهای اندوتلیال زخمها، مهاجرت و تشکیل لوله مویرگی میگردد. ولی بیشترین نقش آن در واسطه شدن تعاملات بین اندوتلیال-پرسیت و پایداری رگ است (43).
8- لپتین: باعث تکثیر سلولهای اندوتلیال و افزایش بیان MMP میشود(44).
9-E-selectin : باعث مهاجرت و تمایز سلولهای اندوتلیال میشود(18).
10- هپارین: باعث تسهیل اتصال VEGF و FGF به رسپتورهای سلولی و افزایش سطح NO میشود(45).
11-Epherin : فعـالکننـده رسپتورهــای تیروزین کینازی
است. این فاکتور در تنظیم توسعه مورفوژنز رگها و همچنین در تمایز شریان از ورید دخالت دارد. به این صورت که EpherinB4سرنوشت رگ را به سمت تشکیل عروق وریدی میبرد در حالی که EpherinB2 سرنوشت را به سمت عروق شریانی میبرد(46).
12- ماکروفاژها: کار آنها هماهنگ و موزون کردن فیوژن رگها و تشکیل آنها است. ماکروفاژها نقش سلولهای پل را بین سلولهای رأسی در حال ملحق شدن به هم بازی میکنند(19، 10).
13- متالوپروتئینازها یا MMP : این فاکتور با شناخته شدن آنها در تکامل قورباغه توسط گراس مطرح شد. MMP به چهار دسته 1-کلاژناز 2-ژلاتیناز 3-استرومیلینازو 4-MMP های غشایی TM MMP)) تقسیم میشوند.MMP اندوپپتیدازهایی هستند که از درون، بر زنجیره پلیپپتیدی اثر کرده و یک یا چند اسیدآمینه را از بخش درونی ساختمان کلاژن قطع میکنند، باعث هضم کلاژن نوع 4 در ساختار ECM میشوند و زمینه را برای مهاجرت و تکثیر سلولهای اندوتلیال فراهم میسازند(47).
14-Ang-Tie2 : رسپتور Tie2در سلولهای اندوتلیال و برخی از سلولهای خونساز در طی تکوین بیان میشود. برای رگزایی طبیعی ضروری است و به سه لیگاند متفاوت Ang1، Ang2 و Ang4 متصل میشود. آنژیوپویتین خودش شامل Ang1 و Ang2 میشود که Ang1-Tie2 جزو محرکها و مسئول پایداری رگها است.
به این صورت که از طریق افزایش چسبندگی EC-EC باعث بقای سلولهای اندوتلیال میشود و توسط فیبروبلاست و سلولهای حمایتکننده عروق و سلولهای سرطانی بیان میشود. این فاکتور هم چنین مهاجرت دسته جمعی سلولهای اندوتلیال را به لبه پیشرو که توسط یک جابهجاگر غیر معمولی PKC انجام میشود، ارتقا میدهد.Ang2-Tie2 نقش دوگانه در رگزایی دارد به طوری که ضمن رشد تومور و افزایش متابولیتهای آن مانند VEGF باعث تحریک رگزایی میشود. Ang2در کل باعث ناپایداری رگها میشود به این صورت که در حضور محرکهای رگزایی منجر به رگزایی گردیده و در صورت عدم حضـور محرکهـای رگزایـی و فاکتورهـای
احیای سلولهای اندوتلیـال، منجـر به پسرفت رگ میگردد(48).
15- اینتگرین: مهمترین رسپتورهای سطح سلول درگیر در تعامل سلول- ماتریکس هستند. اینها رسپتورهای هترودایمریک ترانس ممبران هستند که از زیر واحدهای a و bتشکیل شدهاند. باهم قرار گرفتن سطح فعالی از MMP2 به همراه اینتگرین aVb3 در رگزایی رگهای خونی، بازتابی از همراه شدن حرکت و تخریب ماتریکس برای تهاجم سلولها است. اینتگرینها باعث جذب سلولهای در حال جوانهزدن نیز میشوند. بیان شدن و فعالشدن مکانیسمهای پروتئولیتیکی EC ها از طریق فعالشدن سیگنالهای وابسته به اینتگرین میباشد(20، 7).
16- PDGFβ: برای جذب و تکثیر پرسیتها در طول رگزایی حیاتی هستند و به عنوان میتوژن برای فیبروبلاستها و سلولهای عضلات صاف و دیگر سلولهای مزانشیمال عمل میکنند(49).
17- پرسیتها: جزء محرکها و باعث پایداری رگها از طریق کاهش تکثیر سلولهای اندوتلیال، کاهش بلوغ و نشت عروق، میشوند. تمایز سلولهای پیشساز به پرسیتها از طریق TGFβ صورت میگیرد. پرسیتهای بیانکننده PDGFRβ در پاسخ به PDGFتولید شده از سلولهای اندوتلیال مهاجرت کرده و عروق تازه تشکیل شده را احاطه میکنند. پرسیتها همچنین با بیانS1P ، خواص سـد بـودن سلولهـای اندوتلیال را از طریق افزایش بیان N-cagherin بین EC پرسیت تنظیم میکنند (50).
18- VE-cadherin: مولکولهای اتصالی سلولهای اندوتلیال هستند که برای چسبندگی سلولهای اندوتلیال و احیای آنها ضروری است. باعث پایداری رگها از طریق مهار سیگنالهای VEGFR2 و فعالکردن مسیر TGFR میشود. توسط غلظت اکسیژن تنظیم میگردد. یک کاهش در VE-cadherinسلولهای همسایه از طریق اندوسیتوز در پاسخ به محرکهای رگزایی منجر به پشتیبانی تشکیل عروق جدید میشود. این فاکتور هم چنین در فیلوپدیای سلول رأسی باعث ایجاد یک ارتباط جدید با دیگر جوانههای عروق میگردد(21، 10).
بیشتر تحریک میکنند. سلولهای ساقه همچنین با رسوب غشای پایه و جذب پرسیتها، موجب پایداری رگ درحال تشکیل میشوند. پیشسازهای پرسیت(سلولهای پیشساز مزانشیمی) از طریق بیان PDGF از سلولهای اندوتلیال جذب رگها میشوند و در پاسخ به TGFβ به پرسیتها تمایز مییابند. پرسیتها با کاهش مهاجرت، تکثیر سلولهای اندوتلیال و کاهش نشت عروق منجر به پایداری رگهای نو ظهور میشوند(32).
شرایط لازم و فاکتورهای دخیل در تشکیل لومن و تکثیر ساقه(شکل 2):
آزادسازی فاکتورهای محرک رگزایی از ECM
راهنمایی و چسبندگی سلول رأسی
مهاجرت سلولهای اندوتلیال به هم پیوسته
طویل شدن ساقه
تشکیل لومن
به کارگیری ماکروفاژها و سلولهای میلوئید
استخدام پرسیتها(33، 32)
2- بلوغ به واسطه پایداری سلولهای فالانکس:
هنگامی که ادغام صورت میگیرد، یک لومن متصل به هم برای عبور جریان خون در سرتاسر مویرگ جدید به وجود میآید. عبور جریان خون و دریافت اکسیژن و مواد غذایی توسط بافت هیپوکسی منجر به کاهش سطح سیگنالهای رگزایی، غیر فعال شدن سنسورهای اکسیژن سلولهای اندوتلیال و افزایش مولکولهای محرک پایداری منجر به پایداری سلولهای اندوتلیال میشود(34). سلولهای اندوتلیال دوباره فنوتیپ فالانکس پایدار را به صورت یک تک لایه محکم و مقاوم با یک سطح ساده برای انتقال جریان خون و تنظیم جریان خون به بافتها، از سر میگیرند.
سلولهای اندوتلیال از طریق سیگنالهای پاراکرین و اتوکرین و سلولهای حمایتکننده اطراف از طریق Notch، Ang1، FGF و VEGF از فنوتیپ سلولهای اندوتلیال پایدار نگهداری و از رگها در مقابل شیب استرس حفاظت میکنند. در نهایت با پایداری و بلوغ، رگ سدی را بین خون و بافت اطراف جهت کنترل تبادل مایعات و مواد تشکیل میدهد(35)(شکل 3).
شرایط لازم برای بلوغ رگها:
برقراری دوباره اتصالات سلول به سلول
رسوب غشا پایه
بلوغ پرسیتها
پایداری
ایجاد سلولهای فالانکس(36)
نقش رگزایی در تومور و متاستاز:
سلولهای تومور جمعیتی از سلولهای میزبان هستند که توانایی تنظیم تکثیر خود را از دست دادهاند و بنابراین به مقدار نامحدود تکثیر میشوند. بافت توموری میتواند مواد تغذیهای و اکسیژن کافی را از طریق انتشار ساده تا محدوده 1 تا 2 میلیمتر جذب نماید و از این نقطه به بعد نیازمند ایجاد رگهای تغذیهکننده جدید میباشند. در طی فرآیندی که بسیار به رگزایی نرمال شباهت دارد، یک تومور میتواند تشکیل رگهای جدید را از شبکه مویرگی موجود القا کند(38، 37).
تعدادی از عوامل فعالکننده:
1- VEGF: فاکتور رشد اندوتلیوم به عنوان فاکتور نفوذپذیری عروق نیز شناخته میشود. این فاکتور یک پروتئین همودایمر باند شده به هپارین است که دارای وزنی معادل 45 کیلو دالتون میباشد. VEGF دارای 7 ایزوفرم PIGF ، A ، B ، C ، D ، E و F که نتیجه اسپلایسینگ متفات از ژن VEFG میباشد، است.VEGF از طریق 1- تنظیم ساخت DNA باعث تکثیر سلولهای اندوتلیال 2- افزایش مؤلفههای آنتیآپوپتیک مانند BcL2 و A1 باعث بقا 3- تخریب غشا پایه باعث مهاجرت 4- فسفریله کردن اجزاء چسبنده بین سلولهای اندوتلیال و اتصالات محکم باعث نفوذپذیری سلولهای اندوتلیال میشود(39).
2- رسپتورهای VEGFR: VEGF دارای سه نوع گیرنده تیروزین کینازی به نامهای VEGFR3 (flt4)، VEGFR1 (flt1) و VEGFR2 (KDR/flk1) میباشد که این گیرندهها بر روی سلولهای اندوتلیال عروق وجود دارند.VEGFA بهVEGFR1 و VEGFR2 متصل میشود. VEGFB و PIGF تنها به VEGFR1 متصل میشود وVEGFD و VEGFC محکم به VEGFR3 متصل میشوند.VEGFA با افینیتی بالاتری به VEGFR1 نسبت به VEGFR2 متصل میشود در حالی که فعالیت تیروزین کینازی VEGFR1ده برابر ضعیفتر از VEGFR2است(40).
3- FGF : فاکتور رشد فیبروبلاست است که شامل 9 عضو مجزا میباشد. از مهمترین FGF ها میتوان بهbFGF و aFGF اشاره کرد که به رسپتورهای تیروزین کینازی FGFR1 و FGFR2 متصل میشود. این فاکتور قویترین فاکتور رگزایی و قابل اتصال به هپارین و هپاران سولفات است. این فاکتور هم به صورت ذخیره شده در غشای پایه به عنوان یک منبع ذخیره و هم به صورت افزایش بیان در طول رگزایی دیده میشود. این فاکتور به طور کلی در مهاجرت و تمایز سلولهای اندوتلیال و پروتئولیز خارجسلولی نقش دارد.FGF ها میتوژن اختصاصی سلولهای اندوتلیال بوده که توسط سلولهای توموری و ماکروفاژهای جذب شده ترشح میشوند(41).
4- HGF : فاکتور رشد هپاتوسیت، که در کاهش آپوپتوز سلولهای اندوتلیال و ترمیم ماتریکس خارجسلولی نقش دارد(15).
5- اینترلوکین 8: باعث تکثیر و مهاجرت سلولهای اندوتلیال و تشکیل تیوپ مویرگی میشود(16).
6-TNFα : یک سایتوکاین التهابی است که در ابتدا توسط ماکروفاژهای فعال شده در طول التهاب و پاسخهای ایمنی ترشح میشود. این فاکتور همچنین با مست سلها، سلولهای اندوتلیال، فیبروبلاستها و مونوسیت مرتبط است. TNFαبه طور مستقیم از طریق القای تمایز و به طور غیرمستقیم از طریق تحریک تولید فاکتورهای رگزایی از دیگر سلولها بر روی سلولهای اندوتلیال اثر میگذارد(42).
7- TGFβ: شامل سه ایزوتایپ TGFβ1، TGFβ2و TGFβ3 میباشد. این فاکتور در بدن باعث افزایش رسوب ECM و افزایش بیان رسپتورهای اینتگرین و واسطه تکثیر سلولهای اندوتلیال زخمها، مهاجرت و تشکیل لوله مویرگی میگردد. ولی بیشترین نقش آن در واسطه شدن تعاملات بین اندوتلیال-پرسیت و پایداری رگ است (43).
8- لپتین: باعث تکثیر سلولهای اندوتلیال و افزایش بیان MMP میشود(44).
9-E-selectin : باعث مهاجرت و تمایز سلولهای اندوتلیال میشود(18).
10- هپارین: باعث تسهیل اتصال VEGF و FGF به رسپتورهای سلولی و افزایش سطح NO میشود(45).
11-Epherin : فعـالکننـده رسپتورهــای تیروزین کینازی
است. این فاکتور در تنظیم توسعه مورفوژنز رگها و همچنین در تمایز شریان از ورید دخالت دارد. به این صورت که EpherinB4سرنوشت رگ را به سمت تشکیل عروق وریدی میبرد در حالی که EpherinB2 سرنوشت را به سمت عروق شریانی میبرد(46).
12- ماکروفاژها: کار آنها هماهنگ و موزون کردن فیوژن رگها و تشکیل آنها است. ماکروفاژها نقش سلولهای پل را بین سلولهای رأسی در حال ملحق شدن به هم بازی میکنند(19، 10).
13- متالوپروتئینازها یا MMP : این فاکتور با شناخته شدن آنها در تکامل قورباغه توسط گراس مطرح شد. MMP به چهار دسته 1-کلاژناز 2-ژلاتیناز 3-استرومیلینازو 4-MMP های غشایی TM MMP)) تقسیم میشوند.MMP اندوپپتیدازهایی هستند که از درون، بر زنجیره پلیپپتیدی اثر کرده و یک یا چند اسیدآمینه را از بخش درونی ساختمان کلاژن قطع میکنند، باعث هضم کلاژن نوع 4 در ساختار ECM میشوند و زمینه را برای مهاجرت و تکثیر سلولهای اندوتلیال فراهم میسازند(47).
14-Ang-Tie2 : رسپتور Tie2در سلولهای اندوتلیال و برخی از سلولهای خونساز در طی تکوین بیان میشود. برای رگزایی طبیعی ضروری است و به سه لیگاند متفاوت Ang1، Ang2 و Ang4 متصل میشود. آنژیوپویتین خودش شامل Ang1 و Ang2 میشود که Ang1-Tie2 جزو محرکها و مسئول پایداری رگها است.
به این صورت که از طریق افزایش چسبندگی EC-EC باعث بقای سلولهای اندوتلیال میشود و توسط فیبروبلاست و سلولهای حمایتکننده عروق و سلولهای سرطانی بیان میشود. این فاکتور هم چنین مهاجرت دسته جمعی سلولهای اندوتلیال را به لبه پیشرو که توسط یک جابهجاگر غیر معمولی PKC انجام میشود، ارتقا میدهد.Ang2-Tie2 نقش دوگانه در رگزایی دارد به طوری که ضمن رشد تومور و افزایش متابولیتهای آن مانند VEGF باعث تحریک رگزایی میشود. Ang2در کل باعث ناپایداری رگها میشود به این صورت که در حضور محرکهای رگزایی منجر به رگزایی گردیده و در صورت عدم حضـور محرکهـای رگزایـی و فاکتورهـای
احیای سلولهای اندوتلیـال، منجـر به پسرفت رگ میگردد(48).
15- اینتگرین: مهمترین رسپتورهای سطح سلول درگیر در تعامل سلول- ماتریکس هستند. اینها رسپتورهای هترودایمریک ترانس ممبران هستند که از زیر واحدهای a و bتشکیل شدهاند. باهم قرار گرفتن سطح فعالی از MMP2 به همراه اینتگرین aVb3 در رگزایی رگهای خونی، بازتابی از همراه شدن حرکت و تخریب ماتریکس برای تهاجم سلولها است. اینتگرینها باعث جذب سلولهای در حال جوانهزدن نیز میشوند. بیان شدن و فعالشدن مکانیسمهای پروتئولیتیکی EC ها از طریق فعالشدن سیگنالهای وابسته به اینتگرین میباشد(20، 7).
16- PDGFβ: برای جذب و تکثیر پرسیتها در طول رگزایی حیاتی هستند و به عنوان میتوژن برای فیبروبلاستها و سلولهای عضلات صاف و دیگر سلولهای مزانشیمال عمل میکنند(49).
17- پرسیتها: جزء محرکها و باعث پایداری رگها از طریق کاهش تکثیر سلولهای اندوتلیال، کاهش بلوغ و نشت عروق، میشوند. تمایز سلولهای پیشساز به پرسیتها از طریق TGFβ صورت میگیرد. پرسیتهای بیانکننده PDGFRβ در پاسخ به PDGFتولید شده از سلولهای اندوتلیال مهاجرت کرده و عروق تازه تشکیل شده را احاطه میکنند. پرسیتها همچنین با بیانS1P ، خواص سـد بـودن سلولهـای اندوتلیال را از طریق افزایش بیان N-cagherin بین EC پرسیت تنظیم میکنند (50).
18- VE-cadherin: مولکولهای اتصالی سلولهای اندوتلیال هستند که برای چسبندگی سلولهای اندوتلیال و احیای آنها ضروری است. باعث پایداری رگها از طریق مهار سیگنالهای VEGFR2 و فعالکردن مسیر TGFR میشود. توسط غلظت اکسیژن تنظیم میگردد. یک کاهش در VE-cadherinسلولهای همسایه از طریق اندوسیتوز در پاسخ به محرکهای رگزایی منجر به پشتیبانی تشکیل عروق جدید میشود. این فاکتور هم چنین در فیلوپدیای سلول رأسی باعث ایجاد یک ارتباط جدید با دیگر جوانههای عروق میگردد(21، 10).
شکل 4: درمانهای ضد رگزایی علیه VEGF و رسپتورهایش(13).
تعدادی از عوامل مهارکننده:
تا به امروز تعداد زیادی از فاکتورهای درونزای (Endogenous) مهارکننده رگزایی شناسایی شدهاند که منشاء بسیاری از آنها به طور طبیعی از ماتریکس خارجسلولی میباشد و برخی هم درواقع پروتیینهای غشای پایه هستند. بر این اساس، مهارکنندههای درونزای رگزایی به دو کلاس اصلی شامل مهارکنندههای مشتق از ماتریکس و مهارکنندههای مشتق نشده از ماتریکس تقسیم میشوند(51).
الف ـ مهارکنندههای مشتق از ماتریکس: 1- کانستاتین 2-
ترومبوسپوندین 1 و 2، 3- اندوستاتین 4- اجزای کلاژن و اجزای فیبرونکتین 5- اندورپلین 6- آرستن(51).
ب- مهارکنندههای مشتقنشده از ماتریکس: 1- آنژیواستاتین 2- آنتیترومبین 3- TIMP 4- اینترفرون 5- اینترلوکین 6- وازوستاتین 7- تروپونین 1، 8- اجزای پرولاکتین 9- SFlt-1 (51).
مکانیسم تاثیر مهارکنندههای رگزایی:
مهارکنندههای رگزایی از چند طریق پدیده رگزایی را مورد هدف قرار میدهند:
1- مهار مسیر VEGFشامل الف) مهارکنندههایPDGF وVEGF که گیرندههای تیروزین کیناز مانند Sunitinib، Sorafenib ،PD173074 ،SU6668 ، SU5416 و FGF را مهار میکنند ب) آنتیبادیهای مونوکلونال مهارکننده VEGF و گیرنده آن.
2- مهارکنندههـای مـاتریکـس متـالوپروتئینازهـا مانندBAY ،AG3340 ، .Marimastat
3- مهارکنندههای درونزاد مانند اندواستاتین، آنژیواستاتین،
تومستاتین، ترومبواسپوندین.
4- مـهـارکـننـدههـای کـیـنازهـای درون سلـولـی ماننــد
مهارکنندههای mTOR، مهارکنندههای پروتئین کینازC.
5- آنتاگونیستهای اینتگرین مانند Vitaxin، آنتی اینتگرین آنتیبادی، پپتیدهای مسدود کننده عملکرد α5β1 و ανβ3 اینتگرین یا Cilengitide.
6- سایتوکاینها و کموکاینها مانند فاکتور نکروز تومور، اینترفرون آلفا و بتا و اینترلوکین(52).
در زیر به برخی از این مکانیسمها اشاره شده است(22، 10):
1- مرگ برنامهریزی شده سلول
2- رونویسی فاکتورهای رشد در سلولهای سرطانی
3- انتقال سیگنال در سلولهای اندوتلیال
4- مهاجرت سلولهای اندوتلیال
5- بیان مولکولهای چسبان به نامE-cadherin
بسیاری از ترکیبات ضد رگزایی که اکنون در مرحله آزمایشهای کلینیکی قرار دارند، ترکیبات طبیعی هستند. تولید دارو از فرآوردههای طبیعی به سرعت در حال رشد و توسعه میباشد، این ترکیبات استراتژی بسیار امیدبخش برای شناسایی عوامل ضدرگزایی و ضد سرطان میباشند(53)(شکل 4).
نتیجهگیری
تهاجم و متاستاز از مشخصههای بیولوژیک تومورهای بدخیم و علت عمده عوارض جسمی و مرگ و میر سرطان میباشد. ادامه رشد نئوپلاسم اولیه و متاستاز بستگی به خونرسانی کافی به آن منطقه دارد. فرآیند تشکیل عروق جدید یا همان رگزایی، به تومورها این امکان را میدهد که فراتر از 1-2 میلیمتر مکعب توسعه یابند. به استثنای تومورهای خوشخیم که رگزایی کمی دارند و سرعت رشد آنها کند است، رگزایی ﻳﻚ ﻓﺮآﻳﻨﺪ ﻣﻬﻢ در ﺷﺮاﻳﻂ ﻓﻴﺰﻳﻮﻟﻮژﻳﻜﻲ و پاتولوژیکی است. گسترش سیستم عروقی، احتمال تهاجم سلولهای توموری را از طریق وارد شدن به جریان خون و انتشار به اندامهای دیگر افزایش میدهد. علاوه بر این، نشان داده شده که تشکیل سیستم عروقی در تومورهای بدخیم با قدرت متاستاز تومور رابطه مستقیم دارد. به طور کلی در بافتهای سالم و پایدار (Quiescent) فاکتورهایی که از رگزایی ممانعت میکنند غالب هستند اما در بافتهایی که به سرعت تقسیم میشوند، مولکولهایی که فرآیند رگزایی را تحریک میکنند غلبه دارند. به ﻫﻤﯿﻦ دﻟﯿﻞ ﻣﻬﺎر رگزایی ﺑﻪ ﻋﻨﻮان یﮏ ﻋﺎﻣﻞ کمکﮐﻨﻨﺪه در درﻣﺎن سرطان شناخته میشود. در نتیجه مطالعههای محققان ﺟﻬﺖ تشخیص ﻣﻜﺎﻧﻴﺴﻢ مولکولی و ﻓﺎﻛﺘﻮرﻫﺎی دﺧـﻴﻞ در اﻳﻦ ﻓﺮآﻳﻨﺪ، ﻣﻲﺗﻮاﻧﺪ زﻣﻴﻨﻪﺳﺎز ﺗﻮﺳﻌﻪ راهﻫﺎی درمانی باشد. با توجه به اهمیت رگزایی در تحقیقات مربوط به کشف و شناسایی فاکتورهای آنژیوژنیک و عوامل مهارکننده رگزایی جهت درمان
بیماریهای مختلف از جمله انواعی از تومورها که با رگزایی ارتباط تنگاتنگی داشته و به آن وابسته هستند، روشهـای مهـار رگزایی که با هدف تداخل با این فرآیند مهم جهتگیری نمودهاند، مسیر امیدوار کنندهای برای درمان سرطان محسوب میگردند. از طرف دیگر، ضرورت مطالعههای بیشتر در زمینه فرآوردههای طبیعی ضد رگزایـی و هم چنیـن مسیرهای مولکولی که منجر به مهار
رگزایی میشود، کاملاً محسوس به نظر میرسد.
تشکر و قدردانی
محققان و نویسندگان این مقاله بدینوسیله مراتب امتنان
خود را از همه عزیزانی که ما را در به ثمر رساندن این مقاله یاری نمودند، اعلام میدارند.
ارسال پیام به نویسنده مسئول
| بازنشر اطلاعات | |
 |
این مقاله تحت شرایط Creative Commons Attribution-NonCommercial 4.0 International License قابل بازنشر است. |
